Acyl-protein thioesterase
Acyl-protein thioesterases are enzymes that cleave off lipid modifications on proteins, located on the sulfur atom of cysteine residues linked via a thioester bond.[1] Acyl-protein thioesterases are part of the α/β hydrolase superfamily of proteins and have a conserved catalytic triad.[2] For that reason, acyl-protein thioesterases are also able to hydrolyze oxygen-linked ester bonds.
 Crystal structure of human APT1, PDB code 1fj2. Alpha helices are in red, beta strands in gold, catalytic site residues in black. The 2 different monomers of the dimer are shaded in green and brown. | |||||||||
| Identifiers | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Symbol | Acyl-protein thioesterases (APTs) | ||||||||
| Pfam | PF02230 | ||||||||
| InterPro | IPR029058 | ||||||||
| |||||||||
Function
Acyl-protein thioesterases are involved in the depalmitoylation of proteins, meaning they cleave off palmitoyl modifications on proteins' cysteine residues. Cellular targets include trimeric G-alpha proteins,[3] ion channels[4] and GAP-43.[5] Moreover, human acyl-protein thioesterases 1 and 2 have been identified as major components in controlling the palmitoylation cycle of the oncogene Ras.[6][7] Depalmitoylation of Ras by acyl-protein thioesterases potentially reduces Ras' affinity to endomembranes, allowing it to be palmitoylated again at the Golgi apparatus and to be directed to the plasma membrane. Acyl-protein thioesterases, therefore, are thought to correct potential mislocalization of Ras.
Known enzymes
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Currently fully validated human acyl-protein thioesterases are APT1[8] and APT2[9] which share 66% sequence homology.[10] Additionally there are a handful of putative acyl-protein thioesterases reported, including the ABHD17 enzyme family.[11][12] In the lysosome, PPT1 of the palmitoyl protein thioesterase family has similar enzymatic activity as acyl-protein thioesterases.
Structure
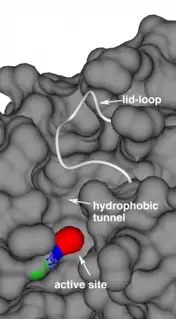
Acyl-protein thioesterases feature 3 major structural components that determine protein function and substrate processing: 1. A conserved, classical catalytic triad to break ester and thioester bonds;[2] 2. A long hydrophobic substrate tunnel to accommodate the palmitoyl moiety, as identified in the crystal structures of human acyl-protein thioesterase 1,[2] human acyl-protein thioesterase 2[13] and Zea mays acyl-protein thioesterase 2;[14] 3. A lid-loop that covers the catalytic site, is highly flexible and is a main factor determining the enzyme's product release rate.[14]

Inhibition
The involvement in controlling the localization of the oncogene Ras has made acyl-protein thioesterases potential cancer drug targets.[15] Inhibition of acyl-protein thioesterases is believed to increase mislocalization of Ras at the cell's membranes, eventually leading to a collapse of the Ras cycle. Inhibitors for acyl-protein thioesterases have been specifically targeting the hydrophobic substrate tunnel,[16][13] the catalytic site serine[17] or both.[18]
Research
Current approaches to study the biological activity of Acyl-protein Thioesterases include proteomics, monitoring the trafficking of microinjected fluorescent substrates,[19][7] the use of cell-permeable substrate mimetics,[20] and cell permeable small molecule fluorescent chemical tools.[21][22][23][24]
References
- Zeidman R, Jackson CS, Magee AI (January 2009). "Protein acyl thioesterases (Review)". Molecular Membrane Biology. 26 (1): 32–41. doi:10.1080/09687680802629329. hdl:10044/1/1452. PMID 19115143.
- Devedjiev Y, Dauter Z, Kuznetsov SR, Jones TL, Derewenda ZS (November 2000). "Crystal structure of the human acyl protein thioesterase I from a single X-ray data set to 1.5 A". Structure. 8 (11): 1137–46. doi:10.1016/s0969-2126(00)00529-3. PMID 11080636.
- Wang A, Yang HC, Friedman P, Johnson CA, Dennis EA (February 1999). "A specific human lysophospholipase: cDNA cloning, tissue distribution and kinetic characterization". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1437 (2): 157–69. doi:10.1016/s1388-1981(99)00012-8. PMID 10064899.
- Tian L, McClafferty H, Knaus HG, Ruth P, Shipston MJ (April 2012). "Distinct acyl protein transferases and thioesterases control surface expression of calcium-activated potassium channels". The Journal of Biological Chemistry. 287 (18): 14718–25. doi:10.1074/jbc.M111.335547. PMC 3340283. PMID 22399288.
- Tomatis VM, Trenchi A, Gomez GA, Daniotti JL (November 2010). "Acyl-protein thioesterase 2 catalyzes the deacylation of peripheral membrane-associated GAP-43". PLOS ONE. 5 (11): e15045. doi:10.1371/journal.pone.0015045. PMC 2994833. PMID 21152083.
- Rocks O, Peyker A, Kahms M, Verveer PJ, Koerner C, Lumbierres M, Kuhlmann J, Waldmann H, Wittinghofer A, Bastiaens PI (March 2005). "An acylation cycle regulates localization and activity of palmitoylated Ras isoforms". Science. 307 (5716): 1746–52. doi:10.1126/science.1105654. PMID 15705808.
- Dekker FJ, Rocks O, Vartak N, Menninger S, Hedberg C, Balamurugan R, Wetzel S, Renner S, Gerauer M, Schölermann B, Rusch M, Kramer JW, Rauh D, Coates GW, Brunsveld L, Bastiaens PI, Waldmann H (June 2010). "Small-molecule inhibition of APT1 affects Ras localization and signaling". Nature Chemical Biology. 6 (6): 449–56. doi:10.1038/nchembio.362. PMID 20418879.
- Duncan JA, Gilman AG (June 1998). "A cytoplasmic acyl-protein thioesterase that removes palmitate from G protein alpha subunits and p21(RAS)". The Journal of Biological Chemistry. 273 (25): 15830–7. doi:10.1074/jbc.273.25.15830. PMID 9624183.
- Tomatis VM, Trenchi A, Gomez GA, Daniotti JL (November 2010). "Acyl-protein thioesterase 2 catalyzes the deacylation of peripheral membrane-associated GAP-43". PLOS ONE. 5 (11): e15045. doi:10.1371/journal.pone.0015045. PMC 2994833. PMID 21152083.
- Conibear E, Davis NG (December 2010). "Palmitoylation and depalmitoylation dynamics at a glance". Journal of Cell Science. 123 (Pt 23): 4007–10. doi:10.1242/jcs.059287. PMC 2987437. PMID 21084560.
- Lin DT, Conibear E (December 2015). "ABHD17 proteins are novel protein depalmitoylases that regulate N-Ras palmitate turnover and subcellular localization". eLife. 4: e11306. doi:10.7554/eLife.11306. PMC 4755737. PMID 26701913.
- Long JZ, Cravatt BF (October 2011). "The metabolic serine hydrolases and their functions in mammalian physiology and disease". Chemical Reviews. 111 (10): 6022–63. doi:10.1021/cr200075y. PMC 3192302. PMID 21696217.
- Won SJ, Davda D, Labby KJ, Hwang SY, Pricer R, Majmudar JD, Armacost KA, Rodriguez LA, Rodriguez CL, Chong FS, Torossian KA, Palakurthi J, Hur ES, Meagher JL, Brooks CL, Stuckey JA, Martin BR (December 2016). "Molecular Mechanism for Isoform-Selective Inhibition of Acyl Protein Thioesterases 1 and 2 (APT1 and APT2)". ACS Chemical Biology. 11 (12): 3374–3382. doi:10.1021/acschembio.6b00720. PMC 5359770. PMID 27748579.
- Bürger M, Willige BC, Chory J (December 2017). "A hydrophobic anchor mechanism defines a deacetylase family that suppresses host response against YopJ effectors". Nature Communications. 8 (1): 2201. doi:10.1038/s41467-017-02347-w. PMC 5736716. PMID 29259199.
- Chavda B, Arnott JA, Planey SL (September 2014). "Targeting protein palmitoylation: selective inhibitors and implications in disease". Expert Opinion on Drug Discovery. 9 (9): 1005–19. doi:10.1517/17460441.2014.933802. PMID 24967607.
- Rusch M, Zimmermann TJ, Bürger M, Dekker FJ, Görmer K, Triola G, Brockmeyer A, Janning P, Böttcher T, Sieber SA, Vetter IR, Hedberg C, Waldmann H (October 2011). "Identification of acyl protein thioesterases 1 and 2 as the cellular targets of the Ras-signaling modulators palmostatin B and M". Angewandte Chemie. 50 (42): 9838–42. doi:10.1002/anie.201102967. PMID 21905186.
- Zimmermann TJ, Bürger M, Tashiro E, Kondoh Y, Martinez NE, Görmer K, Rosin-Steiner S, Shimizu T, Ozaki S, Mikoshiba K, Watanabe N, Hall D, Vetter IR, Osada H, Hedberg C, Waldmann H (January 2013). "Boron-based inhibitors of acyl protein thioesterases 1 and 2". ChemBioChem. 14 (1): 115–22. doi:10.1002/cbic.201200571. PMID 23239555.
- Pedro MP, Vilcaes AA, Tomatis VM, Oliveira RG, Gomez GA, Daniotti JL (2013). "2-Bromopalmitate reduces protein deacylation by inhibition of acyl-protein thioesterase enzymatic activities". PLOS ONE. 8 (10): e75232. doi:10.1371/journal.pone.0075232. PMC 3788759. PMID 24098372.
- Görmer K, Bürger M, Kruijtzer JA, Vetter I, Vartak N, Brunsveld L, Bastiaens PI, Liskamp RM, Triola G, Waldmann H (May 2012). "Chemical-biological exploration of the limits of the Ras de- and repalmitoylating machinery". ChemBioChem. 13 (7): 1017–23. doi:10.1002/cbic.201200078. PMID 22488913.
- Creaser SP, Peterson BR (March 2002). "Sensitive and rapid analysis of protein palmitoylation with a synthetic cell-permeable mimic of SRC oncoproteins". Journal of the American Chemical Society. 124 (11): 2444–5. doi:10.1021/ja017671x. PMID 11890786.
- Kathayat RS, Elvira PD, Dickinson BC (February 2017). "A fluorescent probe for cysteine depalmitoylation reveals dynamic APT signaling". Nature Chemical Biology. 13 (2): 150–152. doi:10.1038/nchembio.2262. PMC 5247352. PMID 27992880.
- Qiu T, Kathayat RS, Cao Y, Beck MW, Dickinson BC (January 2018). "A Fluorescent Probe with Improved Water Solubility Permits the Analysis of Protein S-Depalmitoylation Activity in Live Cells". Biochemistry. 57 (2): 221–225. doi:10.1021/acs.biochem.7b00835. PMC 5823605. PMID 29023093.
- Beck MW, Kathayat RS, Cham CM, Chang EB, Dickinson BC (November 2017). "S-depalmitoylases in live cells and tissues". Chemical Science. 8 (11): 7588–7592. doi:10.1039/C7SC02805A. PMC 5848818. PMID 29568422.
- Kathayat RS, Cao Y, Elvira PD, Sandoz PA, Zaballa ME, Springer MZ, Drake LE, Macleod KF, van der Goot FG, Dickinson BC (January 2018). "Active and dynamic mitochondrial S-depalmitoylation revealed by targeted fluorescent probes". Nature Communications. 9 (1): 334. doi:10.1038/s41467-017-02655-1. PMC 5780395. PMID 29362370.