Alpha-ketoglutarate-dependent hydroxylases
Alpha-ketoglutarate-dependent hydroxylases are a major class of non-heme iron proteins that catalyse a wide range of reactions. These reactions include hydroxylation reactions, demethylations, ring expansions, ring closures, and desaturations.[1][2] Functionally, the αKG-dependent hydroxylases are comparable to cytochrome P450 enzymes. Both use O2 and reducing equivalents as cosubstrates and both generate water.[3]
Biological function
αKG-dependent hydroxylases have diverse roles.[4][5] In microorganisms such as bacteria, αKG-dependent dioxygenases are involved in many biosynthetic pathways.[6][7][8] In plants, αKG-dependent dioxygenases are involved in diverse reactions in plant metabolism.[9] These include flavonoid biosynthesis,[10] and ethylene biosyntheses.[11] In mammals and humans, αKG-dependent dioxygenase have functional roles in biosyntheses (e.g. collagen biosynthesis[12] and L-carnitine biosynthesis[13]), post-translational modifications (e.g. protein hydroxylation[14]), epigenetic regulations (e.g. histone and DNA demethylation[15]), as well as sensors of energy metabolism.[16]
Many αKG-dependent dioxygenase also catalyse uncoupled turnover, in which oxidative decarboxylation of αKG into succinate and carbon dioxide proceeds in the absence of substrate. The catalytic activity of many αKG-dependent dioxygenases are dependent on reducing agents (especially ascorbate) although the exact roles are not understood.[17][18]
Catalytic mechanism
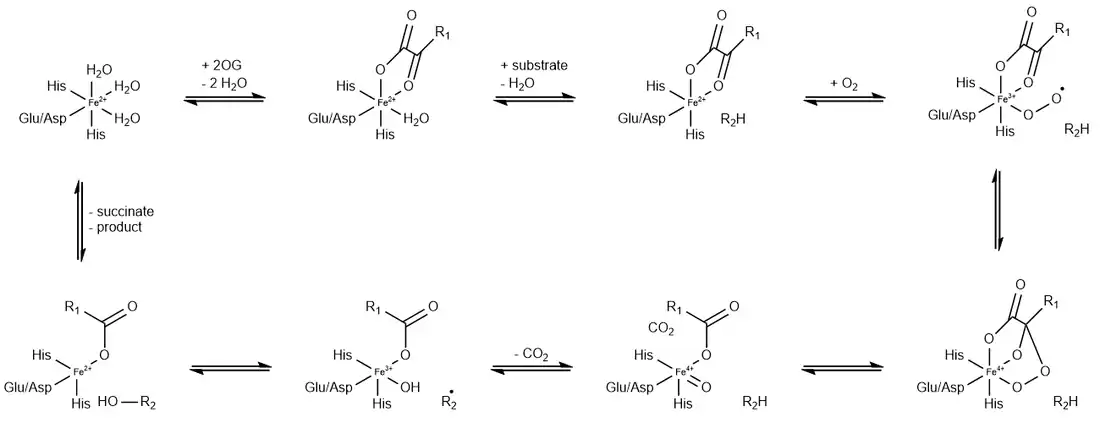
αKG-dependent dioxygenases catalyse oxidation reactions by incorporating a single oxygen atom from molecular oxygen (O2) into their substrates. This conversion is coupled with the oxidation of the cosubstrate αKG into succinate and carbon dioxide.[1][2] With labeled O2 as substrate, the one label appears in the succinate and one in the hydroxylated substrate:[19][20]
- R3CH + O2 + −O2CC(O)CH2CH2CO2− → R3COH + CO2 + −OOCCH2CH2CO2−
The first step involves the binding of αKG and substrate to the active site. αKG coordinates as a bidentate ligand to Fe(II), while the substrate is held by noncovalent forces in close proximity. Subsequently, molecular oxygen binds end-on to Fe cis to the two donors of the αKG. The uncoordinated end of the superoxide ligand attacks the keto carbon, inducing release of CO2 and forming an Fe(IV)-oxo intermediate. This Fe=O center then oxygenates the substrate by an oxygen rebound mechanism.[1][2]
Alternative mechanisms have failed to gain support.[21]
Structure
Protein
All αKG-dependent dioxygenases contain a conserved double-stranded β-helix (DSBH, also known as cupin) fold, which is formed with two β-sheets.[22][23]
Metallocofactor
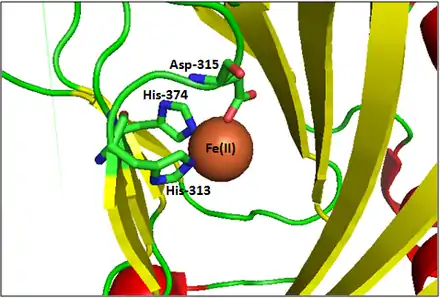
The active site contains a highly conserved 2-His-1-carboxylate (HXD/E...H) amino acid residue triad motif, in which the catalytically-essential Fe(II) is held by two histidine residues and one aspartic acid/glutamic acid residue. The N2O triad binds to one face of the Fe center, leaving three labile sites available on the octahedron for binding αKG and O2.[1][2] A similar facial Fe-binding motif, but featuring his-his-his array, is found in cysteine dioxygenase.
Substrate and cosubstrate binding
The binding of αKG and substrate has been analyzed by X-ray crystallography, molecular dynamics calculations, and NMR spectroscopy. The binding of the ketoglutarate has been observed using enzyme inhibitors.[24]
Some αKG-dependent dioxygenases bind their substrate through an induced fit mechanism. For example, significant protein structural changes have been observed upon substrate binding for human prolyl hydroxylase isoform 2 (PHD2),[25][26][27] a αKG-dependent dioxygenase that is involved in oxygen sensing,[28] and isopenicillin N synthase (IPNS), a microbial αKG-dependent dioxygenase.[29]
Inhibitors
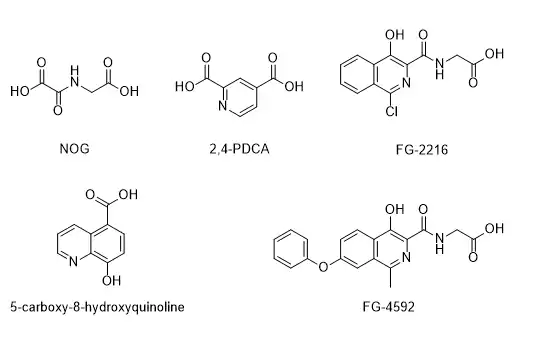
Given the important biological roles that αKG-dependent dioxygenase play, many αKG-dependent dioxygenase inhibitors were developed. The inhibitors that were regularly used to target αKG-dependent dioxygenase include N-oxalylglycine (NOG), pyridine-2,4-dicarboxylic acid (2,4-PDCA), 5-carboxy-8-hydroxyquinoline, FG-2216 and FG-4592, which were all designed mimic the co-substrate αKG and compete against the binding of αKG at the enzyme active site Fe(II).[30][31] Although they are potent inhibitors of αKG-dependent dioxygenase, they lack selectivity and hence sometimes being referred to as so-called 'broad spectrum' inhibitors.[32] Inhibitors that compete against the substrate were also developed, such as peptidyl-based inhibitors that target human prolyl hydroxylase domain 2 (PHD2)[33] and Mildronate, a drug molecule that is commonly used in Russia and Eastern Europe that target gamma-butyrobetaine dioxygenase.[34][35][36]
Assays
Many assays were developed to study αKG-dependent dioxygenases so that information such as enzyme kinetics, enzyme inhibition and ligand binding can be obtained. Nuclear magnetic resonance (NMR) spectroscopy is widely applied to study αKG-dependent dioxygenases.[37] For example, assays were developed to study ligand binding,[38][39][40] enzyme kinetics,[41] modes of inhibition[42] as well as protein conformational change.[43] Mass spectrometry is also widely applied. It can be used to characterise enzyme kinetics,[44] to guide enzyme inhibitor development,[45] study ligand and metal binding[46] as well as analyse protein conformational change.[47] Assays using spectrophotometry were also used,[48] for example those that measure 2OG oxidation,[49] co-product succinate formation[50] or product formation.[51] Other biophysical techniques including (but not limited to) isothermal titration calorimetry (ITC)[52] and electron paramagnetic resonance (EPR) were also applied.[53] Radioactive assays that uses 14C labelled substrates were also developed and used.[54] Given αKG-dependent dioxygenases require oxygen for their catalytic activity, oxygen consumption assay was also applied.[55]
Further reading
- Martinez, Salette; Hausinger, Robert P. (2015-08-21). "Catalytic Mechanisms of Fe(II)- and 2-Oxoglutarate-dependent Oxygenases". The Journal of Biological Chemistry. 290 (34): 20702–20711. doi:10.1074/jbc.R115.648691. ISSN 0021-9258. PMC 4543632. PMID 26152721.
- Hegg EL, Que L Jr (December 1997). "The 2-His-1-carboxylate facial triad--an emerging structural motif in mononuclear non-heme iron(II) enzymes". Eur. J. Biochem. 250 (3): 625–629. doi:10.1111/j.1432-1033.1997.t01-1-00625.x. PMID 9461283..
- Myllylä R, Tuderman L, Kivirikko KI (November 1977). "Mechanism of the prolyl hydroxylase reaction. 2. Kinetic analysis of the reaction sequence". Eur. J. Biochem. 80 (2): 349–357. doi:10.1111/j.1432-1033.1977.tb11889.x. PMID 200425.
- Valegård K, Terwisscha van Scheltinga AC, Dubus A, Ranghino G, Oster LM, Hajdu J, Andersson I (January 2004). "The structural basis of cephalosporin formation in a mononuclear ferrous enzyme" (PDF). Nat. Struct. Mol. Biol. 11 (1): 95–101. doi:10.1038/nsmb712. PMID 14718929. S2CID 1205987.
- Price JC, Barr EW, Tirupati B, Bollinger JM Jr, Krebs C (June 2003). "The first direct characterization of a high-valent iron intermediate in the reaction of an alpha-ketoglutarate-dependent dioxygenase: a high-spin FeIV complex in taurine/alpha-ketoglutarate dioxygenase (TauD) from Escherichia coli". Biochemistry. 42 (24): 7497–7508. doi:10.1021/bi030011f. PMID 12809506.
- Proshlyakov DA, Henshaw TF, Monterosso GR, Ryle MJ, Hausinger RP (February 2004). "Direct detection of oxygen intermediates in the non-heme Fe enzyme taurine/alpha-ketoglutarate dioxygenase". J. Am. Chem. Soc. 126 (4): 1022–1023. doi:10.1021/ja039113j. PMID 14746461.
- Hewitson KS, Granatino N, Welford RW, McDonough MA, Schofield CJ (April 2005). "Oxidation by 2-oxoglutarate oxygenases: non-haem iron systems in catalysis and signalling". Phil. Trans. R. Soc. A. 363 (1829): 807–828. Bibcode:2005RSPTA.363..807H. doi:10.1098/rsta.2004.1540. PMID 15901537. S2CID 8568103.
- Wick CR, Lanig H, Jäger CM, Burzlaff N, Clark T (November 2012). "Structural Insight into the Prolyl Hydroxylase PHD2: A Molecular Dynamics and DFT Study". Eur. J. Inorg. Chem. 2012 (31): 4973–4985. doi:10.1002/ejic.201200391.
References
- Flashman E, Schofield CJ (February 2007). "The most versatile of all reactive intermediates?". Nat. Chem. Biol. 3 (2): 86–87. doi:10.1038/nchembio0207-86. PMID 17235343.
- Hausinger RP (January–February 2004). "Fe(II)/α-ketoglutarate-dependent hydroxylases and related enzymes". Crit. Rev. Biochem. Mol. Biol. 39 (1): 21–68. doi:10.1080/10409230490440541. PMID 15121720. S2CID 85784668.
- Solomon EI, Decker A, Lehnert N (April 2003). "Non-heme iron enzymes: contrasts to heme catalysis". Proc. Natl. Acad. Sci. U.S.A. 100 (7): 3589–3594. doi:10.1073/pnas.0336792100. PMC 152966. PMID 12598659.
- Prescott AG, Lloyd MD (August 2000). "The iron(II) and 2-oxoacid-dependent dioxygenases and their role in metabolism". Nat. Prod. Rep. 17 (4): 367–383. doi:10.1039/A902197C. PMID 11014338.
- Loenarz C, Schofield CJ (January 2011). "Physiological and biochemical aspects of hydroxylations and demethylations catalyzed by human 2-oxoglutarate oxygenases". Trends Biochem. Sci. 36 (1): 7–18. doi:10.1016/j.tibs.2010.07.002. PMID 20728359.
- Scotti JS, Leung IK, Ge W, Bentley MA, Paps J, Kramer HB, Lee J, Aik W, Choi H, Paulsen SM, Bowman LA, Loik ND, Horita S, Ho CH, Kershaw NJ, Tang CM, Claridge TD, Preston GM, McDonough MA, Schofield CJ (September 2014). "Human oxygen sensing may have origins in prokaryotic elongation factor Tu prolyl-hydroxylation". Proc. Natl. Acad. Sci. U.S.A. 111 (37): 13331–13336. Bibcode:2014PNAS..11113331S. doi:10.1073/pnas.1409916111. PMC 4169948. PMID 25197067.
- Clifton IJ, Doan LX, Sleeman MC, Topf M, Suzuki H, Wilmouth RC, Schofield CJ (June 2003). "Crystal structure of carbapenem synthase (CarC)". J. Biol. Chem. 278 (23): 20843–20850. doi:10.1074/jbc.M213054200. PMID 12611886. S2CID 9662423.
- Kershaw NJ, Caines ME, Sleeman MC, Schofield CJ (September 2005). "The enzymology of clavam and carbapenem biosynthesis". Chem. Commun. (34): 4251–4263. doi:10.1039/b505964j. PMID 16113715.
- Farrow SC, Facchini PJ (October 2014). "Functional diversity of 2-oxoglutarate/Fe(II)-dependent dioxygenases in plant metabolism". Front. Plant Sci. 5: 524. doi:10.3389/fpls.2014.00524. PMC 4191161. PMID 25346740.
- Cheng AX, Han XJ, Wu YF, Lou HX (January 2014). "The function and catalysis of 2-oxoglutarate-dependent oxygenases involved in plant flavonoid biosynthesis". Int. J. Mol. Sci. 15 (1): 1080–1095. doi:10.3390/ijms15011080. PMC 3907857. PMID 24434621.
- Zhang Z, Ren JS, Clifton IJ, Schofield CJ (October 2004). "Crystal structure and mechanistic implications of 1-aminocyclopropane-1-carboxylic acid oxidase - the ethylene-forming enzyme". Chem. Biol. 11 (10): 1383–1394. doi:10.1016/j.chembiol.2004.08.012. PMID 15489165.
- Myllyharju J (March 2003). "Prolyl 4-hydroxylases, the key enzymes of collagen biosynthesis". Matrix Biol. 22 (1): 15–24. doi:10.1016/S0945-053X(03)00006-4. PMID 12714038.
- Leung IK, Krojer TJ, Kochan GT, Henry L, von Delft F, Claridge TD, Oppermann U, McDonough MA, Schofield CJ (December 2010). "Structural and mechanistic studies on γ-butyrobetaine hydroxylase". Chem. Biol. 17 (12): 1316–1324. doi:10.1016/j.chembiol.2010.09.016. PMID 21168767.
- Markolovic, Suzana; Wilkins, Sarah E.; Schofield, Christopher J. (2015-08-21). "Protein Hydroxylation Catalyzed by 2-Oxoglutarate-dependent Oxygenases". The Journal of Biological Chemistry. 290 (34): 20712–20722. doi:10.1074/jbc.R115.662627. ISSN 1083-351X. PMC 4543633. PMID 26152730.
- Walport LJ, Hopkinson RJ, Schofield CJ (December 2012). "Mechanisms of human histone and nucleic acid demethylases". Curr. Opin. Chem. Biol. 16 (5–6): 525–534. doi:10.1016/j.cbpa.2012.09.015. PMID 23063108.
- Salminen, A; Kauppinen, A; Kaarniranta, K (2015). "2-Oxoglutarate-dependent dioxygenases are sensors of energy metabolism, oxygen availability, and iron homeostasis: potential role in the regulation of aging process". Cell Mol Life Sci. 72 (20): 3897–914. doi:10.1007/s00018-015-1978-z. PMID 26118662. S2CID 14310267.
- Myllylä R, Majamaa K, Günzler V, Hanauske-Abel HM, Kivirikko KI (May 1984). "Ascorbate is consumed stoichiometrically in the uncoupled reactions catalyzed by prolyl 4-hydroxylase and lysyl hydroxylase". J. Biol. Chem. 259 (9): 5403–5405. doi:10.1016/S0021-9258(18)91023-9. PMID 6325436.
- Flashman E, Davies SL, Yeoh KK, Schofield CJ (March 2010). "Investigating the dependence of the hypoxia-inducible factor hydroxylases (factor inhibiting HIF and prolyl hydroxylase domain 2) on ascorbate and other reducing agents" (PDF). Biochem. J. 427 (1): 135–142. doi:10.1042/BJ20091609. PMID 20055761.
- Welford RW, Kirkpatrick JM, McNeill LA, Puri M, Oldham NJ, Schofield CJ (September 2005). "Incorporation of oxygen into the succinate co-product of iron(II) and 2-oxoglutarate dependent oxygenases from bacteria, plants and humans". FEBS Lett. 579 (23): 5170–5174. doi:10.1016/j.febslet.2005.08.033. hdl:10536/DRO/DU:30019701. PMID 16153644. S2CID 11295236.
- Grzyska PK, Appelman EH, Hausinger RP, Proshlyakov DA (March 2010). "Insight into the mechanism of an iron dioxygenase by resolution of steps following the FeIV=HO species". Proc. Natl. Acad. Sci. U.S.A. 107 (9): 3982–3987. doi:10.1073/pnas.0911565107. PMC 2840172. PMID 20147623.
- Tarhonskaya H, Szöllössi A, Leung IK, Bush JT, Henry L, Chowdhury R, Iqbal A, Claridge TD, Schofield CJ, Flashman E (April 2014). "Studies on deacetoxycephalosporin C synthase support a consensus mechanism for 2-oxoglutarate dependent oxygenases". Biochemistry. 53 (15): 2483–2493. doi:10.1021/bi500086p. PMID 24684493.
- McDonough MA, Loenarz C, Chowdhury R, Clifton IJ, Schofield CJ (December 2010). "Structural studies on human 2-oxoglutarate dependent oxygenases". Curr. Opin. Struct. Biol. 20 (6): 659–672. doi:10.1016/j.sbi.2010.08.006. PMID 20888218.
- Clifton IJ, McDonough MA, Ehrismann D, Kershaw NJ, Granatino N, Schofield CJ (April 2006). "Structural studies on 2-oxoglutarate oxygenases and related double-stranded beta-helix fold proteins". J. Inorg. Biochem. 100 (4): 644–669. doi:10.1016/j.jinorgbio.2006.01.024. PMID 16513174.
- You, Z.; Omura, S.; Ikeda, H.; Cane, D.E.; Jogl, G. (2007). "Crystal structure of the non-heme iron dioxygenase PtlH in pentalenolactone biosynthesis". J. Biol. Chem. 282 (2): 36552–60. doi:10.1074/jbc.M706358200. PMC 3010413. PMID 17942405.
- McDonough MA, Li V, Flashman E, Chowdhury R, Mohr C, Liénard BM, Zondlo J, Oldham NJ, Clifton IJ, Lewis J, McNeill LA, Kurzeja RJ, Hewitson KS, Yang E, Jordan S, Syed RS, Schofield CJ (June 2006). "Cellular oxygen sensing: Crystal structure of hypoxia-inducible factor prolyl hydroxylase (PHD2)". Proc. Natl. Acad. Sci. U.S.A. 103 (26): 9814–9819. Bibcode:2006PNAS..103.9814M. doi:10.1073/pnas.0601283103. PMC 1502536. PMID 16782814.
- Chowdhury R, McDonough MA, Mecinović J, Loenarz C, Flashman E, Hewitson KS, Domene C, Schofield CJ (July 2009). "Structural basis for binding of hypoxia-inducible factor to the oxygen-sensing prolyl hydroxylases". Structure. 17 (7): 981–989. doi:10.1016/j.str.2009.06.002. PMID 19604478.
- Chowdhury R, Leung IK, Tian YM, Abboud MI, Ge W, Domene C, Cantrelle FX, Landrieu I, Hardy AP, Pugh CW, Ratcliffe PJ, Claridge TD, Schofield CJ (August 2016). "Structural basis for oxygen degradation domain selectivity of the HIF prolyl hydroxylases". Nat. Commun. 7: 12673. Bibcode:2016NatCo...712673C. doi:10.1038/ncomms12673. PMC 5007464. PMID 27561929.
- William C, Nicholls L, Ratcliffe P, Pugh C, Maxwell P (2004). "The prolyl hydroxylase enzymes that act as oxygen sensors regulating destruction of hypoxia-inducible factor α". Advan. Enzyme Regul. 44: 75–92. doi:10.1016/j.advenzreg.2003.11.017. PMID 15581484.
- Roach PL, Clifton IJ, Hensgens CM, Shibata N, Schofield CJ, Hajdu J, Baldwin JE (June 1997). "Structure of isopenicillin N synthase complexed with substrate and the mechanism of penicillin formation". Nature. 387 (6635): 827–830. doi:10.1038/42990. PMID 9194566. S2CID 205032251.
- Rose NR, McDonough MA, King ON, Kawamura A, Schofield CJ (August 2011). "Inhibition of 2-oxoglutarate dependent oxygenases". Chem. Soc. Rev. 40 (8): 4364–4397. doi:10.1039/c0cs00203h. PMID 21390379.
- Yeh TL, Leissing TM, Abboud MI, Thinnes CC, Atasoylu O, Holt-Martyn JP, Zhang D, Tumber A, Lippl K, Lohans CT, Leung IK, Morcrette H, Clifton IJ, Claridge TD, Kawamura A, Flashman E, Lu X, Ratcliffe PJ, Chowdhury R, Pugh CW, Schofield CJ (September 2017). "Molecular and cellular mechanisms of HIF prolyl hydroxylase inhibitors in clinical trials". Chem. Sci. 8 (11): 7651–7668. doi:10.1039/C7SC02103H. PMC 5802278. PMID 29435217.
- Hopkinson RJ, Tumber A, Yapp C, Chowdhury R, Aik W, Che KH, Li XS, Kristensen JB, King ON, Chan MC, Yeoh KK, Choi H, Walport LJ, Thinnes CC, Bush JT, Lejeune C, Rydzik AM, Rose NR, Bagg EA, McDonough MA, Krojer T, Yue WW, Ng SS, Olsen L, Brennan PE, Oppermann U, Muller-Knapp S, Klose RJ, Ratcliffe PJ, Schofield CJ, Kawamura A (August 2013). "5-Carboxy-8-hydroxyquinoline is a Broad Spectrum 2-Oxoglutarate Oxygenase Inhibitor which Causes Iron Translocation". Chem. Sci. 4 (8): 3110–3117. doi:10.1039/C3SC51122G. PMC 4678600. PMID 26682036.
- Kwon HS, Choi YK, Kim JW, Park YK, Yang EG, Ahn DR (July 2011). "Inhibition of a prolyl hydroxylase domain (PHD) by substrate analog peptides". Bioorg. Med. Chem. Lett. 21 (14): 4325–4328. doi:10.1016/j.bmcl.2011.05.050. PMID 21665470.
- Sesti C, Simkhovich BZ, Kalvinsh I, Kloner RA (Mar 2006). "Mildronate, a novel fatty acid oxidation inhibitor and antianginal agent, reduces myocardial infarct size without affecting hemodynamics". Journal of Cardiovascular Pharmacology. 47 (3): 493–9. doi:10.1097/01.fjc.0000211732.76668.d2 (inactive 2021-01-10). PMID 16633095.CS1 maint: DOI inactive as of January 2021 (link)
- Liepinsh E, Vilskersts R, Loca D, Kirjanova O, Pugovichs O, Kalvinsh I, Dambrova M (Dec 2006). "Mildronate, an inhibitor of carnitine biosynthesis, induces an increase in gamma-butyrobetaine contents and cardioprotection in isolated rat heart infarction". Journal of Cardiovascular Pharmacology. 48 (6): 314–9. doi:10.1097/01.fjc.0000250077.07702.23. PMID 17204911. S2CID 1812127.
- Hayashi Y, Kirimoto T, Asaka N, Nakano M, Tajima K, Miyake H, Matsuura N (May 2000). "Beneficial effects of MET-88, a gamma-butyrobetaine hydroxylase inhibitor in rats with heart failure following myocardial infarction". European Journal of Pharmacology. 395 (3): 217–24. doi:10.1016/S0014-2999(00)00098-4. PMID 10812052.
- Mbenza NM, Vadakkedath PG, McGillivray DJ, Leung IK (December 2017). "NMR studies of the non-haem Fe(II) and 2-oxoglutarate-dependent oxygenases". J. Inorg. Biochem. 177: 384–394. doi:10.1016/j.jinorgbio.2017.08.032. PMID 28893416.
- Leung IK, Demetriades M, Hardy AP, Lejeune C, Smart TJ, Szöllössi A, Kawamura A, Schofield CJ, Claridge TD (January 2013). "Reporter ligand NMR screening method for 2-oxoglutarate oxygenase inhibitors". J. Med. Chem. 56 (2): 547–555. doi:10.1021/jm301583m. PMC 4673903. PMID 23234607.
- Leung IK, Flashman E, Yeoh KK, Schofield CJ, Claridge TD (January 2010). "Using NMR solvent water relaxation to investigate metalloenzyme-ligand binding interactions". J. Med. Chem. 53 (2): 867–875. doi:10.1021/jm901537q. PMID 20025281.
- Khan A, Leśniak RK, Brem J, Rydzik AM, Choi H, Leung IK, McDonough MA, Schofield CJ, Claridge TD (February 2017). "Development and application of ligand-based NMR screening assays for γ-butyrobetaine hydroxylase". Med. Chem. Commun. 7 (5): 873–880. doi:10.1039/C6MD00004E.
- Hopkinson RJ, Hamed RB, Rose NR, Claridge TD, Schofield CJ (March 2010). "Monitoring the activity of 2-oxoglutarate dependent histone demethylases by NMR spectroscopy: direct observation of formaldehyde". ChemBioChem. 11 (4): 506–510. doi:10.1002/cbic.200900713. PMID 20095001. S2CID 42994868.
- Poppe L, Tegley CM, Li V, Lewis J, Zondlo J, Yang E, Kurzeja RJ, Syed R (November 2009). "Different modes of inhibitor binding to prolyl hydroxylase by combined use of X-ray crystallography and NMR spectroscopy of paramagnetic complexes". J. Am. Chem. Soc. 131 (46): 16654–16655. doi:10.1021/ja907933p. PMID 19886658.
- Bleijlevens B, Shivarattan T, Flashman E, Yang Y, Simpson PJ, Koivisto P, Sedgwick B, Schofield CJ, Matthews SJ (September 2008). "Dynamic states of the DNA repair enzyme AlkB regulate product release". EMBO Rep. 9 (9): 872–877. doi:10.1038/embor.2008.120. PMC 2529343. PMID 18617893.
- Flashman E, Bagg EA, Chowdhury R, Mecinović J, Loenarz C, McDonough MA, Hewitson KS, Schofield CJ (February 2008). "Kinetic rationale for selectivity toward N- and C-terminal oxygen-dependent degradation domain substrates mediated by a loop region of hypoxia-inducible factor prolyl hydroxylases". J. Biol. Chem. 283 (7): 3808–3815. doi:10.1074/jbc.M707411200. PMID 18063574. S2CID 34893579.
- Demetriades M, Leung IK, Chowdhury R, Chan MC, McDonough MA, Yeoh KK, Tian YM, Claridge TD, Ratcliffe PJ, Woon EC, Schofield CJ (July 2012). "Dynamic combinatorial chemistry employing boronic acids/boronate esters leads to potent oxygenase inhibitors". Angew. Chem. Int. Ed. 51 (27): 6672–6675. doi:10.1002/anie.201202000. PMID 22639232.
- Mecinović J, Chowdhury R, Liénard BM, Flashman E, Buck MR, Oldham NJ, Schofield CJ (April 2008). "ESI-MS studies on prolyl hydroxylase domain 2 reveal a new metal binding site". ChemMedChem. 3 (4): 569–572. doi:10.1002/cmdc.200700233. PMID 18058781. S2CID 37628097.
- Stubbs CJ, Loenarz C, Mecinović J, Yeoh KK, Hindley N, Liénard BM, Sobott F, Schofield CJ, Flashman E (May 2009). "Application of a proteolysis/mass spectrometry method for investigating the effects of inhibitors on hydroxylase structure". J. Med. Chem. 52 (9): 2799–2805. doi:10.1021/jm900285r. PMID 19364117.
- Proshlyakov DA, McCracken J, Hausinger RP (April 2016). "Spectroscopic analyses of 2-oxoglutarate-dependent oxygenases: TauD as a case study". J. Biol. Inorg. Chem. 22 (2–3): 367–379. doi:10.1007/s00775-016-1406-3. PMC 5352539. PMID 27812832.
- McNeill LA, Bethge L, Hewitson KS, Schofield CJ (January 2005). "A fluorescence-based assay for 2-oxoglutarate-dependent oxygenases". Anal. Biochem. 336 (1): 125–131. doi:10.1016/j.ab.2004.09.019. PMID 15582567.
- Luo L, Pappalardi MB, Tummino PJ, Copeland RA, Fraser ME, Grzyska PK, Hausinger RP (June 2006). "An assay for Fe(II)/2-oxoglutarate-dependent dioxygenases by enzyme-coupled detection of succinate formation". Anal. Biochem. 353 (1): 69–74. doi:10.1016/j.ab.2006.03.033. PMID 16643838.
- Rydzik AM, Leung IK, Kochan GT, Thalhammer A, Oppermann U, Claridge TD, Schofield CJ (July 2012). "Development and application of a fluoride-detection-based fluorescence assay for γ-butyrobetaine hydroxylase". ChemBioChem. 13 (11): 1559–1563. doi:10.1002/cbic.201200256. PMID 22730246. S2CID 13956474.
- Huang CW, Liu HC, Shen CP, Chen YT, Lee SJ, Lloyd MD, Lee HJ (May 2016). "The different catalytic roles of the metal-binding ligands in human 4-hydroxyphenylpyruvate dioxygenase" (PDF). Biochem. J. 473 (9): 1179–1189. doi:10.1042/BCJ20160146. PMID 26936969.
- Flagg SC, Martin CB, Taabazuing CY, Holmes BE, Knapp MJ (August 2012). "Screening chelating inhibitors of HIF-prolyl hydroxylase domain 2 (PHD2) and factor inhibiting HIF (FIH)". J. Inorg. Biochem. 113: 25–30. doi:10.1016/j.jinorgbio.2012.03.002. PMC 3525482. PMID 22687491.
- Cunliffe CJ, Franklin TJ, Gaskell RM (December 1986). "Assay of prolyl 4-hydroxylase by the chromatographic determination of [14C]succinic acid on ion-exchange minicolumns". Biochem. J. 240 (2): 617–619. doi:10.1042/bj2400617. PMC 1147460. PMID 3028379.
- Ehrismann D, Flashman E, Genn DN, Mathioudakis N, Hewitson KS, Ratcliffe PJ, Schofield CJ (January 2007). "Studies on the activity of the hypoxia-inducible-factor hydroxylases using an oxygen consumption assay". Biochem. J. 401 (1): 227–234. doi:10.1042/BJ20061151. PMC 1698668. PMID 16952279.