MDia1
mDia1 (also known as Dia1, Drf1 for Diaphanous-related formin-1, Diaph1, KIAA4062, p140mDia, mKIAA4062, or D18Wsu154e) is a member of the protein family called the formins and is a Rho effector. It is the mouse version of the diaphanous homolog 1 of Drosophila. mDia1 localizes to cells' mitotic spindle and midbody,[1] plays a role in stress fiber and filopodia formation, phagocytosis, activation of serum response factor, formation of adherens junctions, and it can act as a transcription factor.[2] mDia1 accelerates actin nucleation and elongation by interacting with barbed ends (fast-growing ends) of actin filaments. The gene encoding mDia1 is located on Chromosome 18 of Mus musculus and named Diap1.
| Protein diaphanous homolog 1 | |||||||
|---|---|---|---|---|---|---|---|
 Crystal structure of mDIA1 GBD-FH3 in complex with RhoC-GMPPNP | |||||||
| Identifiers | |||||||
| Symbol | mDia1 | ||||||
| Alt. symbols | DRF1 | ||||||
| PDB | 1z2c | ||||||
| RefSeq | NP_031884.1 | ||||||
| UniProt | o08808 | ||||||
| |||||||
mDia1 is highly homologous to Drosophila diaphanous, regulating the cytokinetic ring during cytokinesis.[3] Homologues in other species are known as well, like the human DIAP1, budding yeast Bni1 or fission yeast Cdc12p.[4]
The gene has been knocked-out in mice.[5]
Structure
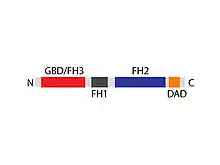
The product of the Diap1 (Diaph1) gene consists of 1255 amino acids resulting in a molecular weight of 139,343 daltons. The mDia1 polypeptide chain can be divided into four protein domains:
- GBD/FH3 (Rho GTPase-binding domain/formin homology 3)domain (366 amino acids long): positions 75-440
- FH1 (formin homology 1) domain (162 amino acids long): positions 586-747
- FH2 (formin homology 2) domain (403 amino acids long): positions 752-1154
- DAD (diaphanous autoregulatory domain) (29 amino acids long): positions 1177-1205
Three supplementary domains were discovered:
- coiled coil (103 amino acids long): positions 460-562
- coiled coil (153 amino acids long): positions 1027-1179
- Arg/Lys-rich domain (4 amino acids long): positions: 1196-1199[3]
The active region of the C terminus consists of formin homology 1 and 2 (FH1 and FH2) and the Dia autoregulatory domain (DAD). The FH1 domain is predicted to be rope-like and it contains binding sites for profilin-actin complexes. The adjacent FH2 domain forms together with the FH2 domain of a second mDia1 molecule a head-to-tail doughnut shaped dimer that encircles the barbed end of an actin filament. Thus the FH2 domain has the ability to dimerize.[2]
The N terminus consists of a Rho GTPase-binding domain (GBD), which is joint to the formin homology 3 (FH3).
DAD can mediate autoinhibition through interactions with the Dia inhibitory domain (DID), which is a subdomain of the GDB/FH3 domain (see section Regulation).
Regulation
Autoinhibition is achieved through binding of the C-terminal DAD to the N-terminal DID. This interaction inhibits the ability of FH2 to nucleate actin assembly.[6] Rho-GTP binds to the GDB domain and disrupts the DAD-DID-interaction thus promoting actin assembly. But this requires high concentrations of Rho-GTP, which may be not physiological. Hence, the release of mDia1 from autoinhibition seems to require nonspecific membrane-associated factors that cooperate with Rho-GTP.[7]
Several binding proteins can regulate mDia1 localization and activity:[2]
- ABI1: Helps to localize mDia1 to lamellipodia, filopodia and cell adhesions
- CLIP 170: Binds the FH2 domain and recruits mDia1 to sites of phagocytosis
- Gα12/13: Helps to localize mDia1 to the leading edge of migrating cells
- RhoA: Required for mDia1 localization to adherens junctions and partially removes mDia's autoinhibition
- RhoB: Helps to localize mDia1 to endosomes
Furthermore the scaffold protein (IQGAP1) seems to impact on mDia1. IQGAP1 regulates the localization of mDia1 to the leading edge to cells. The only tested short C-terminal fragment of IQGAP1 (aa 1503 to 1657) was not activating the mDia1 actin polymerization activity in vitro. However expression of this fragment in macrophages reduced phagocytosis.[8] Thus it remains open if IQGAP1 influences the activity of mDia1 directly like it does for NWASP.[9][10]
Mechanism
Nucleation
In contrast to the Arp 2/3 complex, formins nucleate the formation of unbranched actin filaments. FH2 domains lack structural similarity to actin but can bind actin monomers with very weak affinity.[4] The FH2 dimer nucleates filament assembly by interacting directly with and stabilizing actin polymerization intermediates (dimers and trimers).
Elongation
A formin dimer remains constantly bound to the plus end of an actin filament despite ongoing polymerization. One formin of a dimer dissociates from the barbed end to take the next step while the second formin of the dimer remains bound. Thus the formin dimer processively adds actin monomers to the barbed end and are constantly present at the barbed end of an actin filament (processive capping).[4] The FH1 domain recruits actin monomers through profilin binding, but it does not promote nucleation.[2] Studies demonstrated that FH2 domains protect the rapidly elongating barbed ends of filaments from the vast molar excesses of actin capping proteins.[11][12][13] The precise mechanisms of actin filament nucleation remains an area of active investigation.
The rate of FH2 movement while elongation on an actin filament matches the rate of actin subunit addition, which can exceed 100 subunits per second.
Profilin as a ubiquitous actin-binding protein is associated with most actin monomers in cells. Interactions between profilin-actin with the FH1 domain can accelerate the elongation at the FH2-capped barbed ends.[2]
Function
The formin homology protein mDia1 is a Rho GTPase effector protein, which appears to be universally present in eukaryotic cells and participates in:
- Stress fiber formation
- Endocytosis
- Microtubule functions
Stress fibers are acto-myosin structures, which are important for establishment of cellular tension and thus traction to move a cell ahead; the later is mediated via cell adhesions. Stress fibers are bundles of about 20 actin filaments linked via non-muscle myosin II. In vitro studies showed that mDia1 assembled stress fibers downstream of Rho. Live-cell imaging analysis revealed that mDia1 indeed assembles stress fibers, which are connected at one of their ends to focal adhesions, where actin polymerization occurs.[14] Stress fiber assembly at focal adhesions by mDia1 was later shown to promote their growth and stabilization, suggesting mDia1 exerts effects on the interactions of cells with their environment.[15]
Formins regulate endocytosis. mDia 1 localizes to endosomes and regulates phagocytic cup formation in macrophages.[1]
mDia1 (and mDia2) seems to stabilize microtubules by decreasing the tubulin subunit exchange at their plus ends. The exact mechanism is not fully understood yet. However, the affinity of formins for actin are much higher than for microtubules.[2]
By catalyzing actin polymerization and stabilizing microtubules, mDia1 plays also an important role for cell migration.[16]
Discovery
mDia1 was discovered as p140mDia1 by Watanabe et al.[3] in 1997 as a downstream effector of Rho. A mouse embryo cDNA library was screened to identify a RhoA-GTP-binding protein using a yeast two-hybrid system. Further it was shown that p140mDia1 binds to the GTP-bound form of RhoA only by precipitation from Swiss 3T3 cell lysates. Watanabe et al. could also show the interaction of p140mDia1 with profilin and the colocalization of RhoA, p140mDia and profilin in membrane ruffles of motile cells.
A subsequent study in 1997 by Bione et al.[17] established a link between human DIA and oogenesis, with a defect in the gene leading to premature ovarian failure.
See also
References
- Tominaga T, Sahai E, Chardin P, McCormick F, Courtneidge SA, Alberts AS (2000). "Diaphonous-related fromins bridge Rho GTP-ase and Src tyrosine kinase signaling". Mol. Cell. 5 (1): 13–25. doi:10.1016/S1097-2765(00)80399-8. PMID 10678165.
- Chesarone MA, DuPage AG, Goode BL (2010). "Unleashing Formins to remodel the actin and microtubule cytoskeletons". Nat. Rev. Mol. Cell Biol. 11: 62–74. doi:10.1038/nrm2816. PMID 19997130.
- Watanabe N, Madaule P, Reid T, Ishizaki T, Watanabe G, Kakizuka A, Saito Y, Nakao K, Jockusch BM, Narumiya S (June 1997). "p140mDia, a mammalian homolog of Drosophila diaphanous, is a target protein for Rho small GTPase and is a ligand for profilin". EMBO J. 16 (11): 3044–56. doi:10.1093/emboj/16.11.3044. PMC 1169923. PMID 9214622.
- Goode BL, Eck MJ (2007). "Mechanism and function of formins in the control of actin assembly". Annu. Rev. Biochem. 76: 593–627. doi:10.1146/annurev.biochem.75.103004.142647. PMID 17373907.
- Ercan-Sencicek AG, Jambi S, Franjic D, Nishimura S, Li M, El-Fishawy P, Morgan TM, Sanders SJ, Bilguvar K, Suri M, Johnson MH, Gupta AR, Yuksel Z, Mane S, Grigorenko E, Picciotto M, Alberts AS, Gunel M, Sestan N, State MW (2014). "Homozygous loss of DIAPH1 is a novel cause of microcephaly in humans". European Journal of Human Genetics. 23: 165–72. doi:10.1038/ejhg.2014.82. PMC 4297910. PMID 24781755.
- Nezami AG, Poy F, Eck MJ (February 2006). "Structure of the autoinhibitory switch in formin mDia1". Structure. 14 (2): 257–63. doi:10.1016/j.str.2005.12.003. PMID 16472745.
- Seth A, Otomo C, Rosen MK (August 2006). "Autoinhibition regulates cellular localization and actin assembly activity of the diaphanous-related formins FRLalpha and mDia1". J. Cell Biol. 174 (5): 701–13. doi:10.1083/jcb.200605006. PMC 2064313. PMID 16943183.
- Baarlink C, Grosse R (September 2008). "A GBD uncovered: the FHOD1 N terminus is formin'". Structure. 16 (9): 1287–8. doi:10.1016/j.str.2008.08.002. PMID 18786389.
- Le Clainche C, Schlaepfer D, Ferrari A, Klingauf M, Grohmanova K, Veligodskiy A, Didry D, Le D, Egile C, Carlier MF, Kroschewski R (January 2007). "IQGAP1 stimulates actin assembly through the N-WASP-Arp2/3 pathway". J. Biol. Chem. 282 (1): 426–35. doi:10.1074/jbc.M607711200. PMID 17085436.
- Brandt DT, Marion S, Griffiths G, Watanabe T, Kaibuchi K, Grosse R (July 2007). "Dia1 and IQGAP1 interact in cell migration and phagocytic cup formation". J. Cell Biol. 178 (2): 193–200. doi:10.1083/jcb.200612071. PMC 2064439. PMID 17620407.
- Moseley JB, Sagot I, Manning AL, Xu Y, Eck MJ, Pellman D, Goode BL (2004). "A conserved mechanism for Bni1- and mDia1-induced actin assembly and dual regulation of Bni1 by Bud6 and profilin". Mol. Biol. Cell. 15 (2): 896–907. doi:10.1091/mbc.E03-08-0621. PMC 329402. PMID 14657240.
- Harris ES, Li F, Higgs HN (2004). "The mouse formin, FRLalpha, slows actin filament barbed end elongation, competes with capping protein, accelerates polymerization from monomers, and severs filaments". J. Biol. Chem. 279 (19): 20076–20087. doi:10.1074/jbc.M312718200. PMID 14990563.
- Zigmond SH, Evangelista M, Boone C, Yang C, Dar AC, Sicheri F, Forkey J, Pring M (2003). "Formin leaky cap allows elongation in the presence of tight capping proteins". Curr. Biol. 13 (20): 1820–1823. doi:10.1016/j.cub.2003.09.057. PMID 14561409.
- Hotulainen P, Lappalainen P (2006). "Stress fibers are generated by two distinct actin assembly mechanisms in motile cells". J. Cell Biol. 173 (3): 383–394. doi:10.1083/jcb.200511093. PMC 2063839. PMID 16651381.
- Oakes, Patrick W.; Beckham, Yvonne; Stricker, Jonathan; Gardel, Margaret L. (2012-02-06). "Tension is required but not sufficient for focal adhesion maturation without a stress fiber template". The Journal of Cell Biology. 196 (3): 363–374. doi:10.1083/jcb.201107042. ISSN 1540-8140. PMC 3275371. PMID 22291038.
- Yamana N, Arakawa Y, Nishino T, Kurokawa K, Tanji M, Itoh RE, Monypenny J, Ishizaki T, Bito H, Nozaki K, Hashimoto N, Matsuda M, Narumiya S (2006). "The Rho-mDia1 Pathway Regulates Cell Polarity and Focal Adhesion Turnover in Migrating Cells through Mobilizing Apc and c-Src". Mol. Cell. Biol. 26 (1): 6844–6858. doi:10.1128/MCB.00283-06. PMC 1592856. PMID 16943426.
- Bione S, Sala C, Manzini C, Arrigo G, Zuffardi O, Banfi S, Borsani G, Jonveaux P, Philippe C, Zuccotti M, Ballabio A, Toniolo D (March 1998). "A human homologue of the Drosophila melanogaster diaphanous gene is disrupted in a patient with premature ovarian failure: evidence for conserved function in oogenesis and implications for human sterility". Am. J. Hum. Genet. 62 (3): 533–41. doi:10.1086/301761. PMC 1376955. PMID 9497258.
External links
 Media related to MDia1 at Wikimedia Commons
Media related to MDia1 at Wikimedia Commons- Protein Database