Mononegavirales
Mononegavirales is an order of negative-strand RNA viruses which have nonsegmented genomes. Some common members of the order are Ebola virus, human respiratory syncytial virus, measles virus, mumps virus, Nipah virus, and rabies virus. All of these viruses cause significant disease in humans. Many other important pathogens of nonhuman animals and plants are also in the group. The order includes eleven virus families: Artoviridae, Bornaviridae, Filoviridae, Lispiviridae, Mymonaviridae, Nyamiviridae, Paramyxoviridae, Pneumoviridae, Rhabdoviridae, Sunviridae, and Xinmoviridae.[1]
| Mononegavirales | |
|---|---|
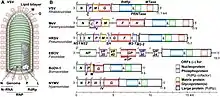 | |
| Vesicular stomatitis virus (VSV) virion and Mononegavirales genomes | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Negarnaviricota |
| Class: | Monjiviricetes |
| Order: | Mononegavirales |
| Families | |
Use of term
The order Mononegavirales (pronounced: /ˌmɒnəˌnɛɡəviˈrɑːlɪz/ MON-ə-NEG-ə-vee-RAH-liz) [note 1][2][3] is a virological taxon that was created in 1991[4][5] and amended in 1995,[6] 1997,[7] 2000,[8] 2005,[9] 2011,[2] 2016,[10] 2017,[11] and 2018.[1] The name Mononegavirales is derived from the Ancient Greek adjective μóνος monos (alluding to the monopartite and single-stranded genomes of most mononegaviruses), the Latin verb negare (alluding to the negative polarity of these genomes), and the taxonomic suffix -virales (denoting a viral order).[3]
Order inclusion criteria
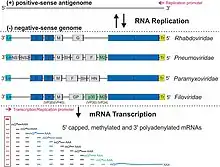
A virus is a member of the order Mononegavirales if[2][3]
- its genome is a linear, typically (but not always) nonsegmented, single-stranded, non-infectious RNA of negative polarity; possesses inverse-complementary 3' and 5' termini; and is not covalently linked to a protein;
- its genome has the characteristic gene order 3'-UTR–core protein genes–envelope protein genes–RNA-dependent RNA polymerase gene–5'-UTR (3'-N-P-M-G-L-5') (there are, however, some exceptions);
- it produces 5–10 distinct mRNAs from its genome via polar sequential transcription from a single promoter located at the 3' end of the genome; mRNAs are 5' capped and polyadenylated;
- it replicates by synthesizing complete antigenomes;
- it forms infectious helical ribonucleocapsids as the templates for the synthesis of mRNAs, antigenomes, and genomes;
- it encodes an RNA-dependent RNA polymerase (RdRp, L) that is highly homologous to those of other mononegaviruses; and/or
- it typically (but not always) produces enveloped virions with a molecular mass of 300–1,000×106; an S20W of 550–>1,045; and a buoyant density in CsCl of 1.18–1.22 g/cm3.
Life cycle
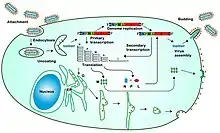
The mononegavirus life cycle begins with virion attachment to specific cell-surface receptors, followed by fusion of the virion envelope with cellular membranes and the concomitant release of the virus nucleocapsid into the cytosol. The virus RdRp partially uncoats the nucleocapsid and transcribes the genes into positive-stranded mRNAs, which are then translated into structural and nonstructural proteins.[9]
Mononegavirus RdRps bind to a single promoter located at the 3' end of the genome. Transcription either terminates after a gene or continues to the next gene downstream. This means that genes close to the 3' end of the genome are transcribed in the greatest abundance, whereas those toward the 5' end are least likely to be transcribed. The gene order is therefore a simple but effective form of transcriptional regulation. The most abundant protein produced is the nucleoprotein, whose concentration in the cell determines when the RdRp switches from gene transcription to genome replication.[9]
Replication results in full-length, positive-stranded antigenomes that are in turn transcribed into negative-stranded virus progeny genome copies. Newly synthesized structural proteins and genomes self-assemble and accumulate near the inside of the cell membrane. Virions bud off from the cell, gaining their envelopes from the cellular membrane they bud from. The mature progeny particles then infect other cells to repeat the cycle.[9]
Paleovirology
Mononegaviruses have a history that dates back several tens of million of years. Mononegavirus "fossils" have been discovered in the form of mononegavirus genes or gene fragments integrated into mammalian genomes. For instance, bornavirus gene "fossils" have been detected in the genomes of bats, fish, hyraxes, marsupials, primates, rodents, ruminants, and elephants.[12][13][14][15][16] Filovirus gene "fossils" have been detected in the genomes of bats, rodents, shrews, tenrecs, and marsupials.[13][14][17][18] A Midway virus "fossil" was found in the genome of zebrafish.[13] Finally, rhabdovirus "fossils" were found in the genomes of crustaceans, mosquitoes, ticks, and plants.[19][14][20][21]
Taxonomy
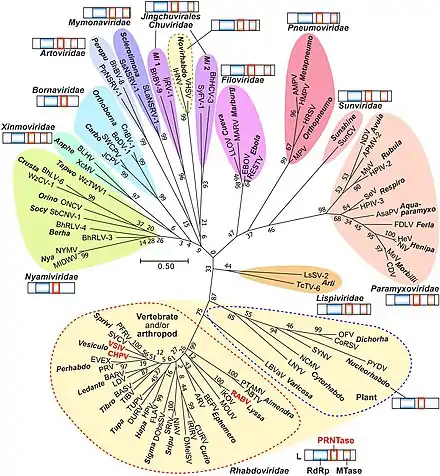
The order has eleven families that include numerous genera, which consist of many different species:
- Artoviridae
- Bornaviridae
- Filoviridae
- Lispiviridae
- Mymonaviridae
- Nyamiviridae
- Paramyxoviridae
- Pneumoviridae
- Rhabdoviridae
- Sunviridae
- Xinmoviridae
Table of the order showing all families, genera, species, and their viruses:[1]
Table legend: "*" denotes type species.
Notes
- According to the rules for taxon naming established by the International Committee on Taxonomy of Viruses (ICTV), the name Mononegavirales is always to be capitalized, italicized, and never abbreviated. The names of the order's physical members ("mononegaviruses" or "mononegavirads") are to be written in lower case, are not italicized, and used without articles.
References
- Amarasinghe GK, Aréchiga Ceballos NG, Banyard AC, Basler CF, Bavari S, Bennett AJ, et al. (April 2018). "Taxonomy of the order Mononegavirales: update 2018". Archives of Virology. 163 (8): 2283–2294. doi:10.1007/s00705-018-3814-x. PMC 6076851. PMID 29637429.
- Easton A, Pringle CR (2011). "Order Mononegavirales". In King AM, Adams MJ, Carstens EB, Lefkowitz EJ (eds.). Virus Taxonomy—Ninth Report of the International Committee on Taxonomy of Viruses. London, UK: Elsevier/Academic Press. pp. 653–657. ISBN 978-0-12-384684-6.
- Kuhn JH, Becker S, Ebihara H, Geisbert TW, Johnson KM, Kawaoka Y, et al. (December 2010). "Proposal for a revised taxonomy of the family Filoviridae: classification, names of taxa and viruses, and virus abbreviations". Archives of Virology. 155 (12): 2083–103. doi:10.1007/s00705-010-0814-x. PMC 3074192. PMID 21046175.
- Pringle, C. R. (1991). "The order Mononegavirales". Archives of Virology. 117 (1–2): 137–40. doi:10.1007/BF01310499. PMID 2006902. S2CID 312908.
- Pringle CR (1991). "Order Mononegavirales". In Francki RI, Fauquet CM, Knudson DK, Brown F (eds.). Classification and Nomenclature of Viruses-Fifth Report of the International Committee on Taxonomy of Viruses. Archives of Virology Supplement, vol. 2. Vienna, Austria: Springer. pp. 239–41. ISBN 978-0-387-82286-0.
- Bishop DH, Pringle CR (1995). "Order Mononegavirales". In Murphy FA, Fauquet CM, Bishop DH, Ghabrial SA, Jarvis AW, Martelli GP, Mayo MA, Summers DM (eds.). Virus Taxonomy—Sixth Report of the International Committee on Taxonomy of Viruses. Archives of Virology Supplement. 10. Vienna, Austria: Springer. pp. 265–267. ISBN 978-3-211-82594-5.
- Pringle CR (1997). "The order Mononegavirales--current status". Archives of Virology. 142 (11): 2321–6. PMID 9672597.
- Pringle CR (2000). "Order Mononegavirales". In van Regenmortel MK, Fauquet CM, Bishop DH, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR, Wickner RB (eds.). Virus Taxonomy—Seventh Report of the International Committee on Taxonomy of Viruses. San Diego, USA: Academic Press. pp. 525–530. ISBN 978-0-12-370200-5.
- Pringle CR (2005). "Order Mononegavirales". In Fauquet CM, Mayo M, Maniloff J, Desselberger U, Ball LA (eds.). Virus Taxonomy—Eighth Report of the International Committee on Taxonomy of Viruses. San Diego, USA: Elsevier/Academic Press. pp. 609–614. ISBN 978-0-12-370200-5.
- Afonso CL, Amarasinghe GK, Bányai K, Bào Y, Basler CF, Bavari S, et al. (August 2016). "Taxonomy of the order Mononegavirales: update 2016". Archives of Virology. 161 (8): 2351–60. doi:10.1007/s00705-016-2880-1. PMC 4947412. PMID 27216929.
- Amarasinghe GK, Bào Y, Basler CF, Bavari S, Beer M, Bejerman N, et al. (August 2017). "Taxonomy of the order Mononegavirales: update 2017". Archives of Virology. 162 (8): 2493–2504. doi:10.1007/s00705-017-3311-7. PMC 5831667. PMID 28389807.
- Horie M, Honda T, Suzuki Y, Kobayashi Y, Daito T, Oshida T, Ikuta K, Jern P, Gojobori T, Coffin JM, Tomonaga K (January 2010). "Endogenous non-retroviral RNA virus elements in mammalian genomes". Nature. 463 (7277): 84–7. Bibcode:2010Natur.463...84H. doi:10.1038/nature08695. PMC 2818285. PMID 20054395.
- Belyi VA, Levine AJ, Skalka AM (July 2010). Buchmeier MJ (ed.). "Unexpected inheritance: multiple integrations of ancient bornavirus and ebolavirus/marburgvirus sequences in vertebrate genomes". PLOS Pathogens. 6 (7): e1001030. doi:10.1371/journal.ppat.1001030. PMC 2912400. PMID 20686665.
- Katzourakis A, Gifford RJ (November 2010). Malik HS (ed.). "Endogenous viral elements in animal genomes". PLOS Genetics. 6 (11): e1001191. doi:10.1371/journal.pgen.1001191. PMC 2987831. PMID 21124940.
- Cui J, Wang LF (November 2015). "Genomic Mining Reveals Deep Evolutionary Relationships between Bornaviruses and Bats". Viruses. 7 (11): 5792–800. doi:10.3390/v7112906. PMC 4664979. PMID 26569285.
- Horie M, Tomonaga K (April 2018). "Paleovirology of bornaviruses: What can be learned from molecular fossils of bornaviruses". Virus Research. 262: 2–9. doi:10.1016/j.virusres.2018.04.006. PMID 29630909.
- Taylor DJ, Leach RW, Bruenn J (June 2010). "Filoviruses are ancient and integrated into mammalian genomes". BMC Evolutionary Biology. 10: 193. doi:10.1186/1471-2148-10-193. PMC 2906475. PMID 20569424.
- Taylor DJ, Dittmar K, Ballinger MJ, Bruenn JA (November 2011). "Evolutionary maintenance of filovirus-like genes in bat genomes". BMC Evolutionary Biology. 11: 336. doi:10.1186/1471-2148-11-336. PMC 3229293. PMID 22093762.
- Metegnier G, Becking T, Chebbi MA, Giraud I, Moumen B, Schaack S, Cordaux R, Gilbert C (2015). "Comparative paleovirological analysis of crustaceans identifies multiple widespread viral groups". Mobile DNA. 6: 16. doi:10.1186/s13100-015-0047-3. PMC 4573495. PMID 26388953.
- Chiba S, Kondo H, Tani A, Saisho D, Sakamoto W, Kanematsu S, Suzuki N (July 2011). Nagy PD (ed.). "Widespread endogenization of genome sequences of non-retroviral RNA viruses into plant genomes". PLOS Pathogens. 7 (7): e1002146. doi:10.1371/journal.ppat.1002146. PMC 3136472. PMID 21779172.
- Fort P, Albertini A, Van-Hua A, Berthomieu A, Roche S, Delsuc F, Pasteur N, Capy P, Gaudin Y, Weill M (January 2012). "Fossil rhabdoviral sequences integrated into arthropod genomes: ontogeny, evolution, and potential functionality". Molecular Biology and Evolution. 29 (1): 381–90. doi:10.1093/molbev/msr226. PMID 21917725.