TBC domain
The TBC (Tre-2/Bub2/Cdc16) is identified as a domain of some proteins or as a protein motif and widely recognized as a conserved one that includes approximately 200 amino acids in all eukaryotes.
Overview
TBC was initially identified as a conserved domain among the tre-2 oncogene product and the yeast cell cycle regulators. It has been shown that humans have almost 42 different TBC proteins which differ from each other by having additional motifs and domains (GRAM, RUN, PTB…) and add functional diversification to the family . The most well known of this protein group are TBC1D1 and TBC1D4 which are directly associated with functional diseases. Moreover, most of them have really close relations with other protein domains. For example, it has been demonstrated that some of them act like a GAP (GTPase-activating protein) for small GTPases: Rab activity is modulated in part by GTPase-activating proteins (GAPs) and many of these RabGAPs share a Tre2/Bub2/Cdc16(TBC)-domain architecture. However, it is needed much research on these kind of proteins and in this article it explains what is known by now.
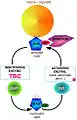
Picture 1. A Rab Cycle in membrane trafficking: The cycle between the GTP-bound inactive state and the GTP- bound active state is led by the Rab protein and regulated by an activating enzyme GEF[1] and an inactivating enzyme GAP which in this case could be the TBC protein. Hereafter, the activated form of Rab, GTP-bound, is incorporated to a specific organelle or vesicle and promotes its transport by interacting with a specific effector molecule. GTPase-activating proteins (GAPs) limit the duration of the active state and accelerate the slow intrinsic rate of GTP hydrolysis.
Functions
TBC mainly functions as a specific Rab GAP (GTPases activating proteins) by being used as tools to inactivate specific membrane trafficking events. GAPs serve to increase GTPase activity by contributing the residues to the active site and promoting conversion from GTP to GDP form. Such activity of TBC proteins does not always require a close physical interaction although few TBC proteins have shown clear GAP activity towards their binding Rabs.[2] Rab families contribute to defining organelles and controlling specificity and rate of transport through individual pathways. Therefore, TBC Rab-GAPS are essential regulators of intracellular and membrane transports as well as central participants in signal transduction. Nevertheless, not all TBC may have a primary role as a Rab-GAP and conversely, not all Rab-GAP contain TBC. In addition, the fact that this family has been poorly studied makes it then further complicated.
Evolution and research
Phylogenetic analysis has provided insight into the evolution of the TBC family. ScrollSaw was implemented as a recent strategy to overcome poor resolution between TBC genes found in standard phylogenetic strategies during initial reconstructions.[3] Significantly, the TBC domain is nearly always smaller than the Rab cohort in any individual genome, suggesting Rab/TBC coevolution. Twenty-one putative TBC sub-classes were founded and identified as a seven robust and two moderately supported clades.
Moreover, there has also been systematic analysis in order to identify the target Rabs of TBC proteins. It was, at first, based on the physical interaction between the TBC domain and its substrate Rab. For instance Barr and his coworkers found a specific interaction between RUTBC3/RabGAP-5 and Rab5A that activates the GTPase activity of Rab5 isoform. Similarly other research has shown that, among other important aspects, the TBC-Rab interaction alone is insufficient to determine the target of TBC proteins. However, there has been a second approach to identifying the target Rabs of TBC by investigating their in vitro GAP activity. Yet there has been similar discrepancies between this findings of different investigators which can be found in literature and may be attributable to differences between methods of in vitro. In addition, research has shown that TBC proteins are associated with some human diseases. For example, a dysfunction of TBC1D1 and TBC1D4 directly affects insulin actions and glucose uptake. Causing overweight or leanness due to the fact that this two family members of TBC regulate insulin-stimulated GLUT4 translocation to the plasma membrane in mammals. Furthermore, many of them have been shown to be associated with cancer, but the exact mechanism by which they are associated with this illness remains largely unknown. Therefore, there’s still much research needed to be done on this biological topic.
References
- Rowlands AG, Panniers R, Henshaw EC (1988). "The catalytic mechanism of guanine nucleotide exchange factor action and competitive inhibition by phosphorylated eukaryotic initiation factor 2". The Journal of Biological Chemistry. 263 (12): 5526–33. PMID 3356695.
- Bos JL, Rehmann H, Wittinghofer A (2007). "GEFs and GAPs: critical elements in the control of small G proteins". Cell. 129 (5): 865–77. doi:10.1016/j.cell.2007.05.018. PMID 17540168. S2CID 15798389.
- Gabernet-Castello C, O'Reilly AJ, Dacks JB, Field MC (2013). "Evolution of Tre-2/Bub2/Cdc16 (TBC) Rab GTPase-activating proteins". Molecular Biology of the Cell. 24 (10): 1574–83. doi:10.1091/mbc.E12-07-0557. PMC 3655817. PMID 23485563.
External links
- Fukuda M (2011). "TBC proteins: GAPs for mammalian small GTPase Rab?". Bioscience Reports. 31 (3): 159–68. doi:10.1042/BSR20100112. PMID 21250943.
- Gabernet-Castello C, O'Reilly AJ, Dacks JB, Field MC (2013). "Evolution of Tre-2/Bub2/Cdc16 (TBC) Rab GTPase-activating proteins". Molecular Biology of the Cell. 24 (10): 1574–83. doi:10.1091/mbc.E12-07-0557. PMC 3655817. PMID 23485563.
- Itoh T, Satoh M, Kanno E, Fukuda M (2006). "Screening for target Rabs of TBC (Tre-2/Bub2/Cdc16) domain-containing proteins based on their Rab-binding activity". Genes to Cells. 11 (9): 1023–37. doi:10.1111/j.1365-2443.2006.00997.x. PMID 16923123.
- Jackson TR, Brown FD, Nie Z, Miura K, Foroni L, Sun J, Hsu VW, Donaldson JG, Randazzo PA (2000). "ACAPs are arf6 GTPase-activating proteins that function in the cell periphery". The Journal of Cell Biology. 151 (3): 627–38. doi:10.1083/jcb.151.3.627. PMC 2185579. PMID 11062263.
- Pan X, Eathiraj S, Munson M, Lambright DG (2006). "TBC-domain GAPs for Rab GTPases accelerate GTP hydrolysis by a dual-finger mechanism". Nature. 442 (7100): 303–6. Bibcode:2006Natur.442..303P. doi:10.1038/nature04847. PMID 16855591. S2CID 4407126.
- Gabernet-Castello C, O'Reilly AJ, Dacks JB, Field MC (2013). "Evolution of Tre-2/Bub2/Cdc16 (TBC) Rab GTPase-activating proteins". Molecular Biology of the Cell. 24 (10): 1574–83. doi:10.1091/mbc.E12-07-0557. PMC 3655817. PMID 23485563.
- Albert S, Will E, Gallwitz D (1999). "Identification of the catalytic domains and their functionally critical arginine residues of two yeast GTPase-activating proteins specific for Ypt/Rab transport GTPases". The EMBO Journal. 18 (19): 5216–25. doi:10.1093/emboj/18.19.5216. PMC 1171592. PMID 10508155.
- Rueckert C, Haucke V (2012). "The oncogenic TBC domain protein USP6/TRE17 regulates cell migration and cytokinesis". Biology of the Cell. 104 (1): 22–33. doi:10.1111/boc.201100108. PMID 22188517. S2CID 5167674.
- Rowlands AG, Panniers R, Henshaw EC (1988). "The catalytic mechanism of guanine nucleotide exchange factor action and competitive inhibition by phosphorylated eukaryotic initiation factor 2". The Journal of Biological Chemistry. 263 (12): 5526–33. PMID 3356695.
- Rangaraju NS, Harris RB (1993). "GAP-releasing enzyme is a member of the pro-hormone convertase family of precursor protein processing enzymes". Life Sciences. 52 (2): 147–53. doi:10.1016/0024-3205(93)90134-o. PMID 8394962.
- Lamarche N, Hall A (1994). "GAPs for rho-related GTPases". Trends in Genetics. 10 (12): 436–40. doi:10.1016/0168-9525(94)90114-7. PMID 7871593.