TMEM221
Transmembrane protein 221 (TMEM221) is a protein that in humans is encoded by the TMEM221 gene.[5] The function of TMEM221 is currently not well understood.
| TMEM221 | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||
| Aliases | TMEM221, transmembrane protein 221 | ||||||||||||||||||||||||
| External IDs | MGI: 3525074 HomoloGene: 110174 GeneCards: TMEM221 | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Orthologs | |||||||||||||||||||||||||
| Species | Human | Mouse | |||||||||||||||||||||||
| Entrez | |||||||||||||||||||||||||
| Ensembl | |||||||||||||||||||||||||
| UniProt | |||||||||||||||||||||||||
| RefSeq (mRNA) | |||||||||||||||||||||||||
| RefSeq (protein) | |||||||||||||||||||||||||
| Location (UCSC) | Chr 19: 17.44 – 17.45 Mb | Chr 8: 71.55 – 71.56 Mb | |||||||||||||||||||||||
| PubMed search | [3] | [4] | |||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||
| |||||||||||||||||||||||||




Gene
General properties
TMEM221 is also known as Putative Transmembrane Protein ENSP00000342162.[6] The TMEM221 gene is 13,159 base pairs long, contains three exons, and is located on the short arm of chromosome 19 at 19p13.11 in humans.[5] It spans from 17,435,509 to 17,448,668 on the minus strand. It is flanked by MVB12A upstream, and by AC010319.5 and NXNL1 downstream.[7]
Promoter
The predicted promoter region (GXP_1485843) is 2016 base pairs long and extends into the beginning of the second exon of TMEM221.[8] The most abundant and highly predicted transcription factors predicted to bind to the promoter are outlined in the table below.
| Transcription Factor | Detailed Matrix Information | Anchor Base | Matrix Similarity | Sequence |
| BRNF | Brn POU domain factors | 11 | 0.905 | caaccatTAATctacttct |
| KLFS | Krueppel like transcription factor | 45 | 0.939 | gggggaatggGGAGtggct |
| LHXF | Lim homeodomain factors | 156 | 0.931 | taaaatgaTTAAttttatgttat |
| HOXF | Paralog hox genes 1-8 from the four hox clusters A, B, C, D | 210 | 0.899 | gcgaaTAATttgggggacc |
| CTCF | CTCF and BORIS gene family, transcriptional regulators with 11 highly conserved zinc finger domains | 256 | 0.813 | cgttgcttcctctaggaGGCTagggag |
| PBXC | Pre B-cell leukemia homeobox 3 | 403 | 1.000 | agcctgagTGACagagc |
| NKRF | Nuclear factor-kappaB repressing factor | 590 | 0.854 | aacTCCTgggc |
| LEFF | T-cell specific HMG-box transcription factor 7 | 701 | 0.879 | actccatCAAAaaaaaa |
| CEBP | Ccaat/Enhancer Binding Protein | 744 | 0.941 | gcagtggtGCAAtct |
| HNFP | Histone nuclear factor P | 763 | 0.843 | ggCGGAggttgcagtgagc |
| CART | Cart-1 (cartilage homeoprotein 1) | 854 | 0.862 | cgggcTAATtttttttttttt |
| TF2B | RNA polymerase II transcription factor II B | 987 | 1.000 | ccgCGCC |
| CAAT | CCAAT binding factor | 1229 | 0.926 | ctggCCAAtatggtg |
| VTBP | Vertebrate TATA binding protein factor | 1342 | 0.852 | tgtttTAAAccacaata |
| PLAG | Pleomorphic adenoma gene | 1419 | 0.844 | ctgtGGGGttcccgcgatccagt |
| NDPK | Nucleoside diphosphate kinase | 1447 | 0.910 | gcAGGGcggggaggccc |
| E2FF | E2F-myc activator/cell cycle regulator | 1471 | 0.989 | gcgggGCGGggcctggc |
| ZF5F | ZF5 POZ domain zinc finger | 1504 | 0.957 | gaggggGCGCccgcg |
| ZF5F | 1505 | 0.957 | ccgcggGCGCcccct | |
| ZTRE | Zinc transcriptional regulatory element | 1525 | 0.988 | ccgGGAGggaggagaaa |
Expression

TMEM221 is highly expressed relative to other human genes in nearly every tissue type, suggesting it could potentially be a housekeeping gene.[9] It is shown consistently to be most highly expressed in the brain, adrenal gland, and ovaries.[10] Conditional expression demonstrates decreased expression of TMEM221 in ovarian cancer, lymphomas, and bone cancer.[11][12][13][14]
mRNA
The longest transcript of TMEM221 is 2,301 base pairs longs. It has one X1 isoform that is 1,547 base pairs long and contains one exon.[5]
Transcript level regulation
The mRNA structure of TMEM221 is predicted to have stem loop formation important for protein recognition and stability. There is one splice enhancer site with two DNAse hypersensitivity sites and bindings sites for transcription factors including CTCF, FOS, NFYB, and NFYA.[7]
Protein
General properties
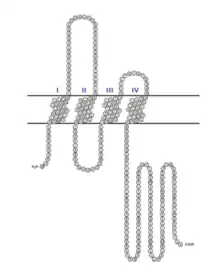
The TMEM221 protein is 291 amino acids long and contains four transmembrane domains. The protein of the X1 variant is 232 amino acids long and contains one transmembrane domain.[16] TMEM221 has a predicted molecular weight of 30 kDa and is slightly basic with a predicted isoelectric point of 8.6.[17]
Composition
TMEM221 has a significantly higher composition of leucine, alanine, and glycine as compared to other human proteins. It also has a significantly lower composition of asparagine and isoleucine. It has no high scoring charge clusters or charged segments.[18]
Domains and Motifs
TMEM221 has two conserved motifs, Jiraiya and DUF5408.[19] The Jiraiya motif was found in all orthologs and composes the three latter transmembrane regions. Jiraiya is reported to be a factor in attenuation of bone morphogenetic protein (BMP) signaling.[20] The DUF5408 motif is not yet characterized.
Structure
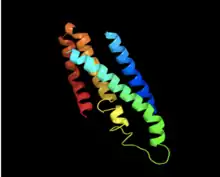
TMEM221 protein is predicted to be composed of approximately 58% random coil, 20% alpha helix, and 22% extended strand.[22][23] The tertiary structure is predicted to have three disulfide bridges between conserved cysteines that are in the non-cytoplasmic regions as well as one crossing between the non-cytoplasmic region and the second transmembrane region.[24]
Subcellular location
TMEM221 is predicted to be mostly localized to the endoplasmic reticulum, but also distributed throughout the mitochondria, vacuolar, plasma membrane, and extracellular space.[25]
Signal peptide
TMEM221 has a predicted signal peptide cleavage site between bases 24 and 25.[26][27]
Post-translational modifications
The only lipid modification TMEM221 has is one palmitoylation site, indicating that its trafficking may not be highly regulated.[28] There is one predicted glycation site.[29] There is one NES signal which is expected as the protein is expected to be located in the cytoplasm, among other locations.[30][31] O-glycosylation is predicted at three sites that are likely important for protein stability and function.[32] There are many possible phosphorylation sites, some with multiple possible kinases, that are likely important for protein activation.[33] There is one SUMOylation site that would aid in nuclear-cytosolic transport.[34]
Homology/evolution
Paralogs
TMEM221 has one predicted paralog, hCG2038292.[6] This protein is mostly highly conserved through the Jiraiya sequence. This diverged approximately 400 million years ago.
Orthologs
TMEM221 is conserved throughout vertebrates but not in invertebrates, plants, or any other organisms. The most distant identified ortholog is the live shark sucker.[35]
| Genus and Species | Common Name | Taxonomic Group | DoD (mya) | Accession Number | Sequence Length (AA) | Percent Identity |
| Homo sapiens | Human | Mammal, Primate | 0 | NP_001177773.1 | 291 | 100% |
| Pan troglodytes | Chimpanzee | Mammal, Primate | 7 | XP_016790933.2 | 291 | 98% |
| Papio anubis | Olive Baboon | Mammal, Primate | 29 | XP_003919243.1 | 290 | 96% |
| Cricetulus griseus | Chinese Hamster | Mammal, Rodentia | 89 | XP_027250594.1 | 291 | 72% |
| Ursus arctos | Grizzly Bear | Mammal, Carnivora | 96 | XP_026356267.1 | 291 | 87% |
| Hipposideros armiger | Great Roundleaf Bat | Mammal, Chiroptera | 96 | XP_019497376.1 | 291 | 85% |
| Trichechus manatus latirostris | West Indian Manatee | Mammal, Sirenia | 105 | XP_004384461.1 | 291 | 86% |
| Phascolarctos cinereus | Koala | Marsupial | 159 | XP_020864777.1 | 288 | 48% |
| Crocodylus porosus | Saltwater Crocodile | Reptile | 312 | XP_019393978.1 | 233 | 38% |
| Egretta garzetta | Little Egret | Bird | 312 | XP_009641067.1 | 209 | 32% |
| Podarcis muralis | Common Wall Lizard | Reptile | 318 | XP_028569471.1 | 305 | 43% |
| Chelonia mydas | Green Sea Turtle | Reptile | 318 | XP_027689962.1 | 280 | 33% |
| Phaethon lepturus | White-tailed Tropicbird | Bird | 318 | XP_010280379.1 | 200 | 30% |
| Python bivittatus | Burmese Python | Reptile | 318 | XP_025027154.1 | 232 | 30% |
| Antrostomus carolinensis | Chuck-will’s-widow | Bird | 318 | XP_010164009.2 | 207 | 23% |
| Xenopus tropicalis | Western Clawed Frog | Amphibian | 352 | XP_004911082.1 | 306 | 37% |
| Xenopus laevis | African Clawed Frog | Amphibian | 352 | NP_001182024.1 | 306 | 37% |
| Danio rerio | Zebrafish | Fish | 433 | XP_003201087.1 | 287 | 37% |
| Salmo trutto | Brown Trout | Fish | 433 | XP_029562717.1 | 304 | 34% |
| Lepisosteus osculatus | Spotted Gard | Fish | 433 | XP_015221220.1 | 170 | 17% |
| Echeneis naucrates | Live Shark Sucker | Fish | 433 | XP_029356055.1 | 312 | 34% |
Rate of Evolution

TMEM221 is a rapidly evolving gene with a rate of divergence faster than cytochrome C, a slowly evolving gene, and fibrinogen, a rapidly evolving gene.
Function/biochemistry
Clinical significance
Disease association
A human disease known to be associated with TMEM221 is amoebiasis, a digestive infection caused by the amoeba Entamoeba histolytica.[6][38] The gene is also shown to be less expressed in a multitude of cancers including ovarian cancer, lymphoma, and bone cancer among others.[12][13][14]
Mutations
There were many potential sites for SNPs in the coding sequence of TMEM221.[39] Notably, W110 has five potential SNPs in its second and third codon positions. There are many other SNPs identified in other conserved amino acids, but these resulted in silent mutations.
| Position in Protein | Mutation Type | Codon Position | Change in Nucleic Acid | Change in Amino Acid | Rs number |
| 13 | Frame Shift/Nonsense | 1 | A → - | M → stop | rs758599058 |
| 33 | Nonsense | 1 | C → T | Q → stop | rs1304244986 |
| 44 | Frame Shift | 3 | G → - | L → C | rs1425689981 |
| 103 | Frame Shift | 3 | G → - | P → L | rs1402509177 |
| 110 | Nonsense
Frameshift Missense Missense Nonsense |
3
3 2 2 2 |
G → A
G → - G → T G → C G → A |
W → stop
W → C W → L W → S W → stop |
rs1448161781
|
| 112 | Missense | 2 | T → C | L → P | rs987960379 |
| 136 | Frame Shift | 3 | A → - | A → H | rs1004328462 |
| 194 | Missense | 1
1 |
G → C
G → A |
D → H
D → N |
rs990835413 |
| 209 | Nonsense | 1 | C → T | Q → stop | rs906917531 |
| 228 | Frame Shift | 1 | G → - | D → T | rs1434673805 |
| 235 | Frame Shift | 3 | + C | T → H | rs149217587 |
| 272 | Missense
Nonsense |
1 | G → A
G → T |
E → K
E → stop |
rs186899872 |
References
- GRCh38: Ensembl release 89: ENSG00000188051 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000043664 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Homo sapiens transmembrane protein 221 (TMEM221), mRNA". 2019-09-12. Cite journal requires
|journal=(help) - "TMEM221 Gene - GeneCards | TM221 Protein | TM221 Antibody". www.genecards.org. Retrieved 2020-05-03.
- "Human hg38 chr19:17,435,509-17,448,668 UCSC Genome Browser v397". genome.ucsc.edu. Retrieved 2020-05-03.
- Genomatix. http://www.genomatix.de/?s=2d8a13e175653babaccb07b76acc8cd2
- "GDS424 / 51886_at". www.ncbi.nlm.nih.gov. Retrieved 2020-05-03.
- Gene NCBI (National Center for Biotechnology Information) https://www.ncbi.nlm.nih.gov/gene/100130519
- "GDS5816 / ILMN_3261382". www.ncbi.nlm.nih.gov. Retrieved 2020-05-03.
- "GDS3754 / 239128_at". www.ncbi.nlm.nih.gov. Retrieved 2020-05-03.
- "GDS5375 / ILMN_3261382". www.ncbi.nlm.nih.gov. Retrieved 2020-05-03.
- "GDS5367 / ILMN_3261382". www.ncbi.nlm.nih.gov. Retrieved 2020-05-03.
- "SOSUI: submit protein sequences". harrier.nagahama-i-bio.ac.jp. Retrieved 2020-05-03.
- NCBI Protein. https://www.ncbi.nlm.nih.gov/protein/300068980
- ExPASy Compute pI/mW tool. https://web.expasy.org/compute_pi/
- SAPS (Stastical Analysis of Protein Sequences). https://www.ebi.ac.uk/Tools/seqstats/saps/
- GenomeNet Motif Search. https://www.genome.jp/tools/motif/
- Aramaki, T., Sasai, N., Yakura, R., Yoshiki, S. (2010). Jiraiya Attenuates BMP Signaling by Interfering with Type II BMP Receptors in Neuroectodermal Patterning. Developmental Cell, 19(4), 547-561. https://doi.org/10.1016/j.devcel.2010.09.001
- Nucleotide NCBI (National Center for Biotechnology Information) https://www.ncbi.nlm.nih.gov/nuccore/NM_001190844.2
- CFSSP (Chou and Fasman Secondary Structure Prediction). http://www.biogem.org/tool/chou-fasman/
- GOR4. https://npsa-prabi.ibcp.fr/cgi-bin/secpred_gor4.pl
- DISULFIND. http://disulfind.dsi.unifi.it/monitor.php?query=v5DhKu
- PSORT II. https://psort.hgc.jp/form2.html
- LipoP. http://www.cbs.dtu.dk/services/LipoP/
- ProP. http://www.cbs.dtu.dk/services/ProP/
- The Cuckoo Workgroup. Prediction of Palmitoylation Sites. http://csspalm.biocuckoo.org/index.php
- NetGlycate. http://www.cbs.dtu.dk/cgi-bin/webface2.fcgi?jobid=5E9CF22B000010DD7A4E9449&wait=20
- NetNES. http://www.cbs.dtu.dk/cgi-bin/webface2.fcgi?jobid=5E9CF2F50000064ECAE3DE03&wait=20
- Phobius. http://phobius.sbc.su.se/cgi-bin/predict.pl
- NetOGlyc. http://www.cbs.dtu.dk/cgi-bin/webface2.fcgi?jobid=5E9CF3370000064E3DFD9AC3&wait=20
- NetPhos. http://www.cbs.dtu.dk/cgi-bin/webface2.fcgi?jobid=5E9CF3BA000010DD71873F80&wait=20
- The Cuckoo Group. Prediction of SUMOylation Sites. http://sumosp.biocuckoo.org/showResult.php
- Blast NCBI (National Center for Biotechnology Information) https://blast.ncbi.nlm.nih.gov/Blast.cgi
- STRING. https://string-db.org/cgi/input.pl?sessionId=nXvhj5eKuvP5&input_page_show_search=on
- Ren, W., Aihara, E., Lei, W. et al. Transcriptome analyses of taste organoids reveal multiple pathways involved in taste cell generation. Sci Rep 7, 4004 (2017). https://doi.org/10.1038/s41598-017-04099-5
- CDC Amebiasis (Center for Disease Control and Prevention) https://www.cdc.gov/parasites/amebiasis/general-info.html
- Gene SNP. https://www.ncbi.nlm.nih.gov/SNP/snp_ref.cgi?locusId=100130519