TRAMP complex
TRAMP complex (Trf4/Air2/Mtr4p Polyadenylation complex) is a multiprotein, heterotrimeric complex having distributive polyadenylation activity and identifies wide varieties of RNAs produced by polymerases.It was originally discovered in Saccharomyces cerevisiae by LaCava et al., Vanacova et al. and Wyers et al. in 2005.[1]
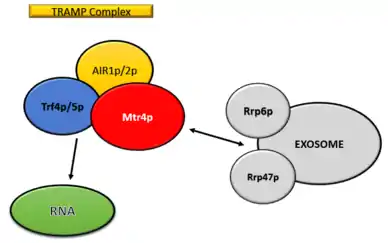
It interacts with the exosome complex in the nucleus of eukaryotic cells and is involved in the 3' end processing and degradation of ribosomal RNA and snoRNAs.[1][2] The TRAMP complex trims the poly(A) tails of RNAs destined for Rrp6 and the core exosome down to 4-5 adenosines assisting in transcript recognition and exosome complex activation.[1][3] The substrate specificity of exosomes is improved in the presence of TRAMP complex as it acts as a crucial cofactor and helps in maintaining various activities.[4]
In this way, TRAMP plays a critical role in ridding the cell of noncoding transcripts generated through pervasive RNA polymerase II transcription, as well as functioning in the biogenesis and turnover of functional coding and noncoding RNAs.[5]
TRAMP complex also affects various other RNA processes either directly or indirectly. It is involved in RNA export, Splicing, hetero-chromatic gene silencing and helps in maintaining stability of genome.[6]
Components
Non-Canonical Poly(A) Polymerases
Pol(A) Polymerases showed various genetic interactions with DNA Topoisomerases Top1p and hence they were called topoisomerase-related function Trf4p and Trf5p[7][8] due to this interaction with DNA it has an important part in genomic stability.[9] In the cell Trf4p is in higher concentration as compared to Trf5p and also has a stronger effect on the phenotype.[10] Trf4p is present throughout the nucleus while Trf5p is mainly found mainly in nucleolus. The Trf4p structure consists of a central domain and a catalytic domain which is similar to the structure of canonical polymerases.[11]
The non-canonical Poly(A) polymerases (Trf4p or Trf5p) of the TRAMP complex which belong to the Cid1 family do not contain RNA recognition motif (RRM) therefore additional proteins like Air1/Air2 are required by the non-canonical polymerases for polyadenylation.[12]
Zinc knuckle proteins
The zinc knuckle proteins Air1p/Air2p (Arginine methyltransferase-interacting RING-finger protein) are mainly involved in the binding of RNAs.[13] There are five CCHC (C stands for Cysteine and H stands for Histidine) zinc knuckle motifs which are present in between the C and N terminals.
In Air2p proteins, the fourth and fifth zinc knuckle have different roles. The fourth zinc knuckle have a role in RNA binding while the fifth knuckle is important for protein-protein interactions.[14] Air2p interacts with the central domain of Trf4p and polyadenylation activity of Trf4p is dependent on this interaction as deletion or mutation of the knuckles hinders the polyadenylation activity.[14] Air1p is responsible for inhibiting methylation of Npl3p (a protein which is responsible mRNA export). Air1p/Air2p also direct abnormal mRNPs to TRAMP pathway and bring about their degradation.[15]
Ski2 like helicase Mtr4p
The Ski2 like helicase Mtr4p was discovered during the screening of heat resistant mutants that gather Poly(A) RNA in the nucleus and is mainly involved in unwinding activity. Mtr4p (also called as Dob1p) is an SF2 helicase and belongs to DExH-box RNA helicases family consisting of two RecA like domains.[16] It also consists of WH domain (Winged Helix domain), an Arch domain (also called as stalk and KOW domain [Kyprides, Ouzounis, Woese domain]) and helical bundle domain.[16] The packing of the WH and helical bundle domains on surface of the helicase core results in the formation of a channel for ssRNA.[16]
Mtr4p requires ATP or dATP hydrolysis for RNA duplex unwinding mediaited by Q-motif. A single-stranded region 3' to the paired region is also essential for the unwinding activity of Mtr4p. Through direct contact with various components of exosome, Mtr4p helps in proper addition of RNA substrates of TRAMP complex to nuclear exosome.[17]
Difference between canonical and non-canonical polymerases
The difference between non-canonical and canonical Poly(A) Polymerases is that canonical polymerases help in maintaining mRNAs and its activity is regulated by a specific sequence in the mRNA[18] while polyadenylation of non-canonical polymerases uses a different regulated sequence in the RNA and specifies RNAs for degeneration or processing.[13] Canonical polymerases belong to DNA polymerase β superfamily whereas non-canonical polymerases belong to Cid1 family, another main difference is the length of the poly(A) tail; canonical polymerases can add many adenylates thus the RNA produced has longer poly(A) tails while non-canonical polymerases on the other hand can produce RNAs with shorter length of poly(A) tails as they can add only few adenylates.[19]
Interaction with 3’->5’ exonuclease complex exosome
The TRAMP complex brings about degradation or processing of various RNAs with the help of 3’->5’ exonuclease complex called the exosome. A hexameric ring of RNase PH (Pleckstrin Homology) domain proteins, Rrp41p, Rrp42p, Rrp43p, Rrp45p, Rrp46p and Mtr3p comprises the exosome of S. cerevisiae.[20] The exosome can bring about RNA degradation more efficiently in the presence of Rrp6p with the help of TRAMP complex invitro. Also, RNA degradation is enhanced in the presence of various exosome cofactors which are recruited co-transcriptionally.[21]
The Ski complex consisting of Ski2p, Ski3p, Ski8p is required by cytoplasmic exosome for all mRNA degradation pathways.[22] The cytoplasmic exosome along with the Ski7p protein attaches to various abnormal ribosomes and mRNAs and brings about their degradation.[20]
Inter-relation between components
All the components of the TRAMP complex are inter-related to each other. For the activity of Poly(A) polymerases likeTrf4p/Trf5p, zinc knuckle proteins are essential. In similar way RNA degradation brought about by exosomes is stimulated by unwinding activity of Ski2 like helicases and Mtr4p which acts as a cofactor. The unwinding activity of Mtr4p is improved by the Trf4p/Air2p in the TRAMP complex.[13] Mtr4p also has an important role in maintaining and controlling the length of Poly(A) tails. But destruction or absence of Mtr4p results in hyperadenylation and hinders the length of Poly(A) tails.
A complex formed between Trf5p, Air1p and Mtr4p is called as TRAMP5 complex.[15] In S. cerevisiae there are two types of TRAMP complexes depending on the presence of polymerases. If Trf4p is present, then the complex is called as TRAMP4 and if Trf5p is present then It is called as TRAMP5.[23]
RNA substrates
RNAs produced by all three polymerases (Pol I, II, III) act as substrates for TRAMP complex. TRAMP complex is involved in processing and surveillance of various RNAs and degrade abnormal RNAs. Different type of RNA substrates include ribosomal RNAs (rRNAs), small nucleolar RNAs (snoRNAs), transfer RNAs (tRNAs), small nuclear RNAs(snRNAs), Long transcripts of RNA polymerase II (Pol II) etc. But the mechanism by which TRAMP complex identifies various substrates is unknown.
The TRAMP complex works more efficiently in RNA processing by engaging Exosome complex exonuclease RrP6 wherein Nab3(RNA binding protein) plays a crucial role.[15][23]
Role in maintaining chromatin
Post-transcriptional modifications due to various enzymes like methyltransferase Hmt1p (Rmt1p) may have an indirect effect on chromatin maintenance. The chromatin structures are affected when RNA substrates of TRAMP complex are transcribed across the genome. Various TRAMP components interact physically and genetically with various proteins and bring about changes in chromatin and DNA metabolism.[1]
Conservation of TRAMP-mediated processes
Components of the TRAMP complex in Saccharomyces cerevisiae are conserved in other organisms ranging from yeast to mammals.The TRAMP complex components of Schizosaccharomyces pombe including Cid14p, Air1p, and Mtr4p are functionally similar to the components of TRAMP complex in S. cerevisiae.[24]
In humans
The TRAMP complex in humans consists of various components including the helicase hMtr4p, a non-canonical poly(A) polymerase hPAPD (PAP-associated domain-containing) 5 or hPAPD7, and a Zinc knuckle protein hZCCHC7, RNA binding protein hRbm7p.[25]
References
- Jia, Huijue; Wang, Xuying; Liu, Fei; Guenther, Ulf-Peter; Srinivasan, Sukanya; Anderson, James T.; Jankowsky, Eckhard (2011-06-10). "The RNA helicase Mtr4p modulates polyadenylation in the TRAMP complex". Cell. 145 (6): 890–901. doi:10.1016/j.cell.2011.05.010. ISSN 1097-4172. PMC 3115544. PMID 21663793.
- Vanácová, Stepánka; Wolf, Jeannette; Martin, Georges; Blank, Diana; Dettwiler, Sabine; Friedlein, Arno; Langen, Hanno; Keith, Gérard; Keller, Walter (June 2005). "A new yeast poly(A) polymerase complex involved in RNA quality control". PLOS Biology. 3 (6): e189. doi:10.1371/journal.pbio.0030189. ISSN 1545-7885. PMC 1079787. PMID 15828860.
- Callahan, Kevin P.; Butler, J. Scott (2010-02-05). "TRAMP complex enhances RNA degradation by the nuclear exosome component Rrp6". The Journal of Biological Chemistry. 285 (6): 3540–3547. doi:10.1074/jbc.M109.058396. ISSN 1083-351X. PMC 2823493. PMID 19955569.
- Schmidt, Karyn; Butler, J. Scott (March 2013). "Nuclear RNA surveillance: role of TRAMP in controlling exosome specificity". Wiley Interdisciplinary Reviews: RNA. 4 (2): 217–231. doi:10.1002/wrna.1155. ISSN 1757-7012. PMC 3578152. PMID 23417976.
- Ciais, Delphine; Bohnsack, Markus T.; Tollervey, David (May 2008). "The mRNA encoding the yeast ARE-binding protein Cth2 is generated by a novel 3' processing pathway". Nucleic Acids Research. 36 (9): 3075–3084. doi:10.1093/nar/gkn160. ISSN 1362-4962. PMC 2396412. PMID 18400782.
- Genome integrity : facets and perspectives. Lankenau, Dirk-Henner. Berlin: Springer. 2007. ISBN 9783540375319. OCLC 164366985.CS1 maint: others (link)
- Sadoff, B. U.; Heath-Pagliuso, S.; Castaño, I. B.; Zhu, Y.; Kieff, F. S.; Christman, M. F. (October 1995). "Isolation of mutants of Saccharomyces cerevisiae requiring DNA topoisomerase I". Genetics. 141 (2): 465–479. ISSN 0016-6731. PMC 1206748. PMID 8647385.
- Castaño, I. B.; Heath-Pagliuso, S.; Sadoff, B. U.; Fitzhugh, D. J.; Christman, M. F. (1996-06-15). "A novel family of TRF (DNA topoisomerase I-related function) genes required for proper nuclear segregation". Nucleic Acids Research. 24 (12): 2404–2410. doi:10.1093/nar/24.12.2404. ISSN 0305-1048. PMC 145947. PMID 8710513.
- Wang, Z.; Castaño, I. B.; De Las Peñas, A.; Adams, C.; Christman, M. F. (2000-08-04). "Pol kappa: A DNA polymerase required for sister chromatid cohesion". Science. 289 (5480): 774–779. doi:10.1126/science.289.5480.774. ISSN 0036-8075. PMID 10926539.
- Reis, Clara C.; Campbell, Judith L. (March 2007). "Contribution of Trf4/5 and the nuclear exosome to genome stability through regulation of histone mRNA levels in Saccharomyces cerevisiae". Genetics. 175 (3): 993–1010. doi:10.1534/genetics.106.065987. ISSN 0016-6731. PMC 1840065. PMID 17179095.
- Huh, Won-Ki; Falvo, James V.; Gerke, Luke C.; Carroll, Adam S.; Howson, Russell W.; Weissman, Jonathan S.; O'Shea, Erin K. (2003-10-16). "Global analysis of protein localization in budding yeast". Nature. 425 (6959): 686–691. doi:10.1038/nature02026. ISSN 1476-4687. PMID 14562095. S2CID 669199.
- Stevenson, Abigail L.; Norbury, Chris J. (2006-10-15). "The Cid1 family of non-canonical poly(A) polymerases". Yeast (Chichester, England). 23 (13): 991–1000. doi:10.1002/yea.1408. ISSN 0749-503X. PMID 17072891.
- Houseley, Jonathan; Tollervey, David (2009-02-20). "The many pathways of RNA degradation". Cell. 136 (4): 763–776. doi:10.1016/j.cell.2009.01.019. ISSN 1097-4172. PMID 19239894. S2CID 17570967.
- Hamill, Stephanie; Wolin, Sandra L.; Reinisch, Karin M. (2010-08-24). "Structure and function of the polymerase core of TRAMP, a RNA surveillance complex". Proceedings of the National Academy of Sciences of the United States of America. 107 (34): 15045–15050. doi:10.1073/pnas.1003505107. ISSN 1091-6490. PMC 2930566. PMID 20696927.
- Anderson, James T.; Wang, Xuying (January 2009). "Nuclear RNA surveillance: no sign of substrates tailing off". Critical Reviews in Biochemistry and Molecular Biology. 44 (1): 16–24. doi:10.1080/10409230802640218. ISSN 1549-7798. PMID 19280429. S2CID 86059909.
- Jackson, Ryan N.; Klauer, A. Alejandra; Hintze, Bradley J.; Robinson, Howard; van Hoof, Ambro; Johnson, Sean J. (2010-07-07). "The crystal structure of Mtr4 reveals a novel arch domain required for rRNA processing". The EMBO Journal. 29 (13): 2205–2216. doi:10.1038/emboj.2010.107. ISSN 1460-2075. PMC 2905245. PMID 20512111.
- Bernstein, Jade; Ballin, Jeff D.; Patterson, Dimeka N.; Wilson, Gerald M.; Toth, Eric A. (2010-12-14). "Unique properties of the Mtr4p-poly(A) complex suggest a role in substrate targeting". Biochemistry. 49 (49): 10357–10370. doi:10.1021/bi101518x. ISSN 1520-4995. PMC 2999651. PMID 21058657.
- Wilusz, Jeremy E.; Spector, David L. (February 2010). "An unexpected ending: noncanonical 3' end processing mechanisms". RNA. 16 (2): 259–266. doi:10.1261/rna.1907510. ISSN 1469-9001. PMC 2811654. PMID 20007330.
- Grzechnik, Pawel; Kufel, Joanna (2008-10-24). "Polyadenylation linked to transcription termination directs the processing of snoRNA precursors in yeast". Molecular Cell. 32 (2): 247–258. doi:10.1016/j.molcel.2008.10.003. ISSN 1097-4164. PMC 2593888. PMID 18951092.
- Houseley, Jonathan; Tollervey, David (April 2008). "The nuclear RNA surveillance machinery: The link between ncRNAs and genome structure in budding yeast?". Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms. Novel RNA nucleotidyl transferases and gene regulation. 1779 (4): 239–246. doi:10.1016/j.bbagrm.2007.12.008. PMID 18211833.
- Schneider, Claudia; Anderson, James T.; Tollervey, David (2007-07-20). "The Exosome Subunit Rrp44 Plays a Direct Role in RNA Substrate Recognition". Molecular Cell. 27 (2): 324–331. doi:10.1016/j.molcel.2007.06.006. ISSN 1097-2765. PMID 17643380.
- Brown, J. T.; Bai, X.; Johnson, A. W. (March 2000). "The yeast antiviral proteins Ski2p, Ski3p, and Ski8p exist as a complex in vivo". RNA. 6 (3): 449–457. doi:10.1017/s1355838200991787. ISSN 1355-8382. PMC 1369926. PMID 10744028.
- Houseley, Jonathan; LaCava, John; Tollervey, David (2006-07-01). "RNA-quality control by the exosome". Nature Reviews Molecular Cell Biology. 7 (7): 529–539. doi:10.1038/nrm1964. ISSN 1471-0080. PMID 16829983. S2CID 22499032.
- Keller, C.; Woolcock, K.; Hess, D.; Buhler, M. (2010). "Proteomic and functional analysis of the noncanonical poly(A) polymerase Cid14". RNA. 16 (6): 1124–1129. doi:10.1261/rna.2053710. PMC 2874164. PMID 20403971.
- Lubas, Michal; Christensen, Marianne S.; Kristiansen, Maiken S.; Domanski, Michal; Falkenby, Lasse G.; Lykke-Andersen, Søren; Andersen, Jens S.; Dziembowski, Andrzej; Jensen, Torben Heick (2011-08-19). "Interaction profiling identifies the human nuclear exosome targeting complex" (PDF). Molecular Cell. 43 (4): 624–637. doi:10.1016/j.molcel.2011.06.028. ISSN 1097-4164. PMID 21855801.