Dinosaur Park Formation
The Dinosaur Park Formation is the uppermost member of the Belly River Group (also known as the Judith River Group), a major geologic unit in southern Alberta. It was deposited during the Campanian stage of the Late Cretaceous, between about 76.9 and 75.8 million years ago.[3] It was deposited in alluvial and coastal plain environments, and it is bounded by the nonmarine Oldman Formation below it and the marine Bearpaw Formation above it.[4]
| Dinosaur Park Formation Stratigraphic range: Campanian, 76.9–75.8 Ma | |
|---|---|
 Dinosaur Park Formation exposed along the Red Deer River in Dinosaur Provincial Park, southeastern Alberta, Canada. | |
| Type | Geological formation |
| Unit of | Belly River Group |
| Underlies | Bearpaw Formation |
| Overlies | Oldman Formation |
| Lithology | |
| Primary | Sandstone (lower) Mudstone and siltstone (upper) |
| Other | Bentonite and coal |
| Location | |
| Coordinates | 49.2°N 110.4°W |
| Approximate paleocoordinates | 56.4°N 75.8°W |
| Region | |
| Country | |
| Extent | Western Canadian Sedimentary Basin |
| Type section | |
| Named for | Dinosaur Provincial Park |
| Named by | Eberth, D.A. and Hamblin, A.P.[1][2] |
| Year defined | 1993 |
 Dinosaur Park Formation (Canada)  Dinosaur Park Formation (Alberta) | |
The Dinosaur Park Formation contains dense concentrations of dinosaur skeletons, both articulated and disarticulated, which are often found with preserved remains of soft tissues. Remains of other animals such as fish, turtles, and crocodilians, as well as plant remains, are also abundant.[5] The formation has been named after Dinosaur Provincial Park, a UNESCO World Heritage Site where the formation is well exposed in the badlands that flank the Red Deer River.[2]
Geological setting

The Dinosaur Park Formation is composed of sediments that were derived from the erosion of the mountains to the west. It was deposited on an alluvial to coastal plain by river systems that flowed eastward and southeastward to the Bearpaw Sea, a large inland sea that was part of the Western Interior Seaway. That sea gradually inundated the adjacent coastal plain, depositing the marine shales of the Bearpaw Formation on top of the Dinosaur Park Formation.[4]
The Dinosaur Park Formation is about 70 metres (230 ft) thick at Dinosaur Park. The lower portion of the formation was laid down in fluvial channel environments and consists primarily of fine- to medium-grained, crossbedded sandstones. The upper portion, which was deposited in overbank and floodplain environments, consists primarily of massive to laminated, organic-rich mudstones with abundant root traces, and thin beds of bentonite. The Lethbridge Coal Zone, which consists of several seams of low-rank coal interbedded with mudstones and siltstones, marks the top of the formation.[4]
The sediments of the Dinosaur Park Formation are similar to those of the underlying Oldman Formation and they were originally included in that formation. The two formations are separated by a regional disconformity, however, and are distinguished by petrographic and sedimentologic differences. In addition, articulated skeletal remains and bonebeds are rare in the Oldman Formation but abundant in the Dinosaur Park Formation.[2][4]
Biostratigraphy
The Dinosaur Park Formation can be divided into at least two distinct faunas. The lower part of the formation is characterized by the abundance of Corythosaurus and Centrosaurus. This group of species is replaced higher in the formation by a different ornithischian fauna characterized by the presence of Lambeosaurus and Styracosaurus. The appearance of several new, rare species of ornithischian at the very top of the formation may indicate that a third distinct fauna had replaced the second during the transition into younger, non-Dinosaur Park sediments, at the same time an inland sea transgresses onto land, but there are fewer remains here. An unnamed pachyrhinosaur, Vagaceratops irvinensis, and Lambeosaurus magnicristatus may be more common in this third fauna.[6][7]
The timeline below follows a synthesis presented by Fowler (2016)[3] with additional information from Arbour et al. 2009,[8] Evans et al. 2009, and Penkalski, 2013.[9] Megaherbivore Assemblage Zones (MAZ) follow data presented by Mallon et al., 2012.[10]

Amphibians
Remains of the following amphibians have been found in the formation:[11]
Albanerpetontidae (extinct, salamander-like amphibians)
- Habrosaurus prodilatus
- Lisserpeton
- Opisthotriton kayi
- Scapherpeton tectum
- unnamed caudatan
- 2 indeterminate caudatans
Salienta (frogs)
Dinosaurs
Remains of the following dinosaurs have been found in the formation:[8][14]
Ornithischians
Remains of the following ornithischians have been found in the formation:[15]
Ankylosaurs
| Ankylosaurs from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Anodontosaurus | A. inceptus | An ankylosaurine ankylosaurid | ||||
| Dyoplosaurus | D. acutosquameus | Lower, 76.5Ma ago[8] | An ankylosaurine ankylosaurid |  | ||
| Edmontonia | E. rugosidens | Lower, 76.5-75.9Ma ago[8] | A nodosaurine nodosaurid |  | ||
| Euoplocephalus | E. tutus | Lower to Middle, ~76.4-75.6Ma[9] | An ankylosaurine ankylosaurid |  | ||
| Panoplosaurus | P. mirus | Middle, 75.6Ma ago[8] | "Partial skeleton with complete skull, osteoderms, additional isolated teeth, postcranial elements, osteoderms."[16] | A nodosaurine nodosaurid |  | |
| Platypelta | P. coombsi | Lower, 77.5-76.5 MA[17] | An ankylosaurine ankylosaurid |  | ||
| Scolosaurus | S. cutleri | Lower, 76.5Ma ago or more[9] | An ankylosaurine ankylosaurid briefly thought to be synonymous with Euoplocephalus. It possibly came from the upper layers of the underlying Oldman Formation.[18] |  | ||
| S. thronus | An ankylosaurine ankylosaurid | |||||
Ceratopsians
An unnamed Pachyrhinosaurus-like taxon has been recovered from the formation.
| Ceratopsians from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Centrosaurus | C. apertus | Middle, 76.2-75.5Ma ago[8] | "[Fifteen] skulls, several skeletons, all adult; abundant bone-bed material with rare juveniles and subadults."[19] C. nasicornis may be a synonym. | A centrosaurine ceratopsid |  | |
| Chasmosaurus | C. belli | Middle, 76-75.5Ma ago[8] | "[Twelve] skulls, several skeletons."[19] | A chasmosaurine ceratopsid |  | |
| C. russelli | Lower, 76.5-76Ma ago[8] | "[Six] complete or partial skulls."[20] | ||||
| Mercuriceratops | M. gemini[21] | Lower, ~77Ma ago[21] | "one apomorphic squamosal"[21] | A chasmosaurine ceratopsid |  | |
| Monoclonius | M. lowei | A dubious centrosaurine ceratopsid. Possibly synonymous with Centrosaurus. | ||||
| Pentaceratops[22] | P. aquilonius[22] | Uppermost, 74.8 MA[22] | two frill fragments[22] | A dubious chasmosaurine ceratopsid that may be the same species as Spiclypeus shipporum.[23] |  | |
| Spinops[24] | S. sternbergorum[24] | Lower, 76.5Ma[24] | "partial parietal bone, partial dentary, unidentifiable limb fragments, partial skull, and partial right squamosal."[24] | A centrosaurine ceratopsid.It may actually be from the upper Oldman Formation.[24] |  | |
| Styracosaurus | S. albertensis | Upper, 75.5-75.2Ma ago[8] | "[Two] skulls, [three] skeletons, additional material in bone beds."[19] | A centrosaurine ceratopsid |  | |
| Unescoceratops | U. koppelhusae | Partial lower jaw[25] | A leptoceratopsid thought to have been between one and two meters long and less than 91 kilograms. Its teeth were the roundest of all leptoceratopsids. | |||
| Vagaceratops | V. irvinensis | Upper, 75Ma ago[8] | "[Three] skulls, skeleton lacking tail."[20] | A chasmosaurine ceratopsid species previously classified as a species of Chasmosaurus.[26] |  | |
Ornithopods
At least one indeterminate hypsilophodont specimen has been recovered from the formation.
In a 2001 review of hadrosaur eggshell and hatchling material from the Dinosaur Park Formation, Darren H. Tanke and M. K. Brett-Surman concluded that hadrosaurs nested in both the ancient upland and lowlands of the formation's depositional environment.[27] The upland nesting grounds may have been preferred by the less common hadrosaurs, like Brachylophosaurus or Parasaurolophus. However, the authors were unable to determine what specific factors shaped nesting ground choice in the formation's hadrosaurs. They suggested that behavior, diet, soil condition, and competition between dinosaur species all potentially influenced where hadrosaurs nested.[28]
Sub-centimeter fragments of pebbly-textured hadrosaur eggshell have been reported from the Dinosaur Park Formation. This eggshell is similar to the hadrosaur eggshell of Devil's Coulee in southern Alberta as well as that of the Two Medicine and Judith River Formations in Montana, United States.[29] While present, dinosaur eggshell is very rare in the Dinosaur Park Formation and is only found in two different microfossil sites.[27] These sites are distinguished by large numbers of pisidiid clams and other less common shelled invertebrates like unionid clams and snails. This association is not a coincidence as the invertebrate shells would have slowly dissolved and released enough basic calcium carbonate to protect the eggshells from naturally occurring acids that otherwise would have dissolved them and prevented fossilization.[29]
In contrast with eggshell fossils, the remains of very young hadrosaurs are actually somewhat common. Darren Tanke has observed that an experienced collector could actually discover multiple juvenile hadrosaur specimens in a single day. The most common remains of young hadrosaurs in the Dinosaur Park Formation are dentaries, bones from limbs and feet, as well as vertebral centra. The material showed little or none of the abrasion that would have resulted from transport, meaning the fossils were buried near their point of origin.[30] Bonebeds 23, 28, 47, and 50 are productive sources of young hadrosaur remains in the formation, especially bonebed 50. The bones of juvenile hadrosaurs and fossil eggshell fragments are not known to have preserved in association with each other, despite both being present in the formation.[31]
| Ornithopods from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Corythosaurus | C. casuarius | Lower-Middle, 76.5-75.5Ma ago[8] | "Approximately [ten] articulated skulls and associated postcrania, [ten to fifteen] articulated skulls, isolated skull elements, juvenile to adult."[32] | A lambeosaurin lambeosaurine hadrosaur |  | |
| Gryposaurus | G. notabilis | Lower, 76.2-76Ma ago[8] | "Approximately [ten] complete skulls, [twelve] fragmentary skulls, associated postcrania."[33] | A kritosaurin saurolophine hadrosaur |  | |
| Lambeosaurus | L. lambei | Upper, 75.5-75Ma ago[8] | "Approximately [seven] articulated skulls with associated postcrania, [possibly ten] articulated skulls, isolated skull elements, juvenile to adult."[34] | |||
| L. magnicristatus | Upper/Bearpaw Formation, 74.8Ma ago[8] | "[Two] complete skulls, one with associated, articulated postcrania."[34] |  | |||
| Parasaurolophus | P. walkeri | Lower, 76.5-75.3Ma ago[7] | "Complete skull and postcranial skeleton."[34] | A parasaurolophin lambeosaurine hadrosaur. |  | |
| Prosaurolophus | P. maximus | Upper, 75.5 – 74.8 Ma | "[Twenty to twenty-five] individuals, including at least [seven] articulated skulls and associated postcrania."[33] | A saurolophin saurolophine hadrosaur |  | |
Pachycephalosaurs
| Pachycephalosaurs from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Foraminacephale | F. brevis | Also present in the Oldman Formation | Frontoparetal dome, various other skull fragments including juvenile and subadult material | Once thought to be a species of Stegoceras |  | |
| Gravitholus | G. albertae | "Frontoparietal dome."[35] |  | |||
| Hanssuesia | H. sternbergi | Lower, also present in the Oldman Formation and Judith River Formation |  | |||
| Stegoceras | S. validum | Specimens including frontoparietal dome.[35] |  | |||
Theropods
In the Dinosaur Park Formation, small theropods are rare due to the tendency of their thin-walled bones to be broken or poorly preserved.[36] Small bones of small theropods that were preyed upon by larger ones may have been swallowed whole and digested.[37] In this context, the discovery of a small theropod dinosaur with preserved tooth marks was especially valuable.[36] Possible indeterminate avimimid remains are known from the formation.
Ornithomimids
| Ornithomimids from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Ornithomimus | O. sp.[38] | Type specimen | An ornithomimid, possibly a species of Struthiomimus.[39] |  | ||
| Qiupalong | Q. sp.[40] | Several specimens | An ornithomimid, possibly the radiation of this genus into Asia.[40] |  | ||
| Rativates | R. evadens | Type specimen | An ornithomimid, formerly a specimen of Struthiomimus.[41] |  | ||
Oviraptorosaurs
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Oviraptorosaurs from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Caenagnathus | C. collinsi | Mandible, type specimen | A caenagnathid[42] which rivalled Anzu in size.[43] |  | ||
| Chirostenotes | C. pergracilis | Several fragmentary specimens, type specimen | A mid-sized caenagnathid. |  | ||
| Citipes | C. elegans[43] | Several fragmentary specimens, type specimen | Smallest caenagnathid from the formation.[43] |  | ||
| Macrophalangia | M. canadensis | Junior synonym of Chirostenotes pergracilis | ||||
Paravians
A new taxon of troodontid based solely on teeth is known from the upper part of the formation.[44]
| Paravians from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| cf. Baptornis | Indeterminate | A hesperornithine bird | ||||
| cf. Cimolopteryx | Indeterminate | Partial coracoid | A possible charadriiform bird | |||
| Dromaeosaurus | D. albertensis | Several specimens and teeth, type specimen | A dromaeosaurid |  | ||
| Hesperonychus | H. elizabethae | Hip bones and partial toes and claws, type specimen | A microraptorine dromaeosaur, also found in the Oldman Formation |  | ||
| Latenivenatrix | L. mcmasterae | Hip bones, pelvis, skull fragments, type specimen | A large troodontid measuring 3–3.5 m (9.8–11.5 ft). | .png.webp) | ||
| cf. Palintropus | Unnamed | Partial shoulder girdles | An ambiortiform bird | |||
| cf. Paronychodon | cf. P. lacustris | Teeth | An indeterminate maniraptoran, also found in the Judith River | |||
| cf. Pectinodon[45] | Indeterminate | Teeth | A troodont | |||
| Polyodontosaurus | P. grandis | Dentary, type specimen | Nomen dubium. Possibly synonymous with Latenivenatrix. | |||
| Richardoestesia | R. gilmorei | Mandible, type specimen | A dromaeosaurid | |||
| R. isosceles[44] | Teeth | |||||
| Saurornitholestes | S. langstoni | Incomplete skeleton and teeth, type specimen. A dentary referred to Saurornitholestes was discovered that preserved tooth marks left by a young tyrannosaur.[46] | A dromaeosaurid |  | ||
| Stenonychosaurus | S. inequalis | Nearly complete skeleton and other partial skeletons, type specimen | A troodontid once thought to be a species of Troodon |  | ||
Tyrannosaurs
| Tyrannosaurs from the Dinosaur Park Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
| Daspletosaurus | Unnamed species[47] | Middle-Upper, 75.6-75Ma ago[8] | Several specimens | A tyrannosaurine tyrannosaurid, also present in the Bearpaw Formation |  | |
| Gorgosaurus | G. libratus | Lower-Middle, 76.6-75.1Ma ago[8] | Numerous specimens, type specimen[47] | An albertosaurine tyrannosaurid |  | |
Color key
|
Notes Uncertain or tentative taxa are in small text; |
Other reptiles
Choristoderes
Choristoderes, or champsosaurs, were aquatic reptiles. Small examples looked like lizards, while larger types were superficially similar to crocodilians. Remains of the following Choristoderes have been found in the formation:[48]
- Champsosaurus (at least 3 species)
- Cteniogenys
Crocodylians
Remains of the following Crocodylians have been found in the formation:[49]
- Albertochampsa
- Leidyosuchus
- at least 1 unnamed taxon
Lizards
Remains of the following lizards have been found in the formation:[50]
- Helodermatids
- Labrodioctes
- Necrosaurids
- Parasaniwa
- Teiids
- Glyptogenys
- Socognathus
- Varanids
- Xenosaurids
- ?Exostinus
Plesiosaurs
Remains of the following Plesiosaurs have been found in the formation:[51]
- indeterminate elasmosaurids (very long-necked) (more common)
- indeterminate polycotylids (shorter-necked)
Pterosaurs
Remains of the following pterosaurs have been found in the formation:[52]
- Cryodrakon [53] (known from small and large specimens)
- 1 unnamed non-azhdarchid pterosaur
Turtles
Remains of the following turtles have been found in the formation:[54]
- Adocus
- "Apalone"
- Aspideretoides (3 species)
- Basilemys
- Boremys
- Judithemys
- Neurankylus
- Plesiobaena
- 2 indeterminate taxa
Mammals
Remains of the following mammals have been found in the formation:[55]
- Multituberculata
- Cimexomys sp.
- Cimolodon spp.
- Cimolomys clarki
- Meniscoessus major
- Mesodma primaeva
- unnamed multituberculates
- Metatherians
- Eutherians
- Cimolestes sp. (uncertain taxonomy)
- Gypsonictops lewisi
- Paranyctoides sternbergi
- Unknown therians: at least 1 species
Fish
Remains of the following fish have been found in the formation:[56]
- Chondrichthyans
- Meristodonoides montanensis (a shark)
- Cretorectolobus olsoni (a carpet shark)
- Eucrossorhinus microcuspidatus (a carpet shark)
- Myledaphus bipartitus (a ray)
- Ischyrhiza mira (a sclerorhynchid)[57]
- Protoplatyrhina renae (a guitarfish)
- indeterminate orectolobid
- Acipenseriformes (sturgeons)
- Holostean fish
- Belonostomus longirostris
- Lepisosteus occidentalis (the gar)
- unnamed bowfin
- at least 2 other holosteans
- Teleost fish
- Paratarpon apogerontus (an elopomorph, like the tarpon)
- Cretophareodus (an osteoglossomorph)
- Coriops amnicolus
- Estesesox foxi
- Oldmanesox
- Paralbula (including Phyllodus)
- at least 8 other teleosts
Invertebrates
Remains of the following invertebrates have been found in the formation:[59]
- Freshwater bivalves
- Freshwater gastropods
- Campeloma (2 species)
- Elimia
- Goniobasis (3 species)
- Hydrobia
- Lioplacodes (2 species)
Flora
Plant body fossils
The following plant body fossils have been found in the formation:[60]
- various ferns
- Equisetum (Equisetaceae)
- Gymnosperms
- Platyspiroxylon (Cupressaceae)
- Podocarpoxylon (Podocarpaceae)
- Elatocladus (Taxodiaceae)
- Sequoia (Taxodiaceae)
- Sequoiaxylon (Taxodiaceae)
- Taxodioxylon (Taxodiaceae)
- Ginkgos
- Angiosperms
- Artocarpus (Moraceae)
- Cercidiphyllum (Cercidiphyllaceae)
- Dombeyopsis (Sterculiaceae)
- Menispermites (Menispermaceae)
- Pistia (Araceae)
- Platanus (Platanaceae)
- Vitis (Vitaceae)
- Trapa (Trapaceae)
Palynomorphs
Palynomorphs are organic-walled microfossils, like spores, pollen, and algae. The following palynomorphs have been found in the formation:[61]
- Unknown producers
- at least 8 species
- Chlorophyta (green algae and blue-green algae)
- at least 12 species
- Pyrrhophyta (dinoflagellates, a type of marine algae)
- unassigned cysts
- Bryophytes (mosses, liverworts, and hornworts)
- Anthocerotophyta (hornworts)
- at least 5 species
- Marchantiophyta (liverworts)
- at least 14 species
- Bryophyta (mosses)
- at least 5 species
- Anthocerotophyta (hornworts)
- Lycopodiophyta
- Lycopodiaceae (club mosses)
- at least 11 species
- Selaginellaceae (small club mosses)
- at least 6 species
- Isoetaceae (quillworts)
- at least 1 species
- Lycopodiaceae (club mosses)
- Polypodiophyta
- Osmundaceae (cinnamon ferns)
- at least 6 species
- Schizaeaceae (climbing ferns)
- at least 20 species
- Gleicheniaceae (Gleichenia and allies; coral ferns)
- at least 5 species
- Cyatheaceae (Cyathea and allies)
- at least 4 species
- Dicksoniaceae (Dicksonia and allies)
- at least 3 species
- Polypodiaceae (ferns)
- at least 4 species
- Matoniaceae
- at least 1 species
- Marsileaceae
- at least 1 species
- Osmundaceae (cinnamon ferns)
- Pinophyta (gymnosperms)
- Cycadaceae (cycads)
- at least 3 species
- Caytoniaceae
- at least 1 species
- Pinaceae (pines)
- at least 4 species
- Cupressaceae (cypresses)
- at least 3 species
- Podocarpaceae (Podocarpus and allies)
- at least 4 species
- Cheirolepidiaceae
- at least 2 species
- Ephedraceae (Mormon teas)
- at least 6 species
- Unknown gymnosperms: at least 3 species
- Cycadaceae (cycads)
- Magnoliophyta (angiosperms)
- Magnoliopsida (dicots)
- Buxaceae (boxwood)
- at least 1 species
- Gunneraceae (gunneras)
- at least 1 species
- Salicaceae (willows, cottonwood, quaking aspen)
- at least 1 species
- Droseraceae (sundews)
- at least 1 species
- Olacaceae (tallowwood)
- at least 2 species
- Loranthaceae (showy mistletoes)
- at least 1 species
- Sapindaceae (soapberry)
- at least 1 species
- Aceraceae (maples)
- at least 1 species
- Proteaceae (proteas)
- at least 9 species
- Compositae (sunflowers)
- at least 1 species
- Fagaceae (beeches, oaks, chestnuts)
- at least 2 species
- Betulaceae (birches, alders)
- at least 1 species
- Ulmaceae (elms)
- at least 1 species
- Chenopodiaceae (goosefoots)
- at least 1 species
- Buxaceae (boxwood)
- Liliopsida (monocots)
- Liliaceae (lilies)
- at least 6 species
- Cyperaceae (sedges)
- at least 1 species
- Sparganiaceae (bur-reeds)
- possibly 1 species
- Unknown angiosperms: at least 88 species
- Liliaceae (lilies)
- Magnoliopsida (dicots)
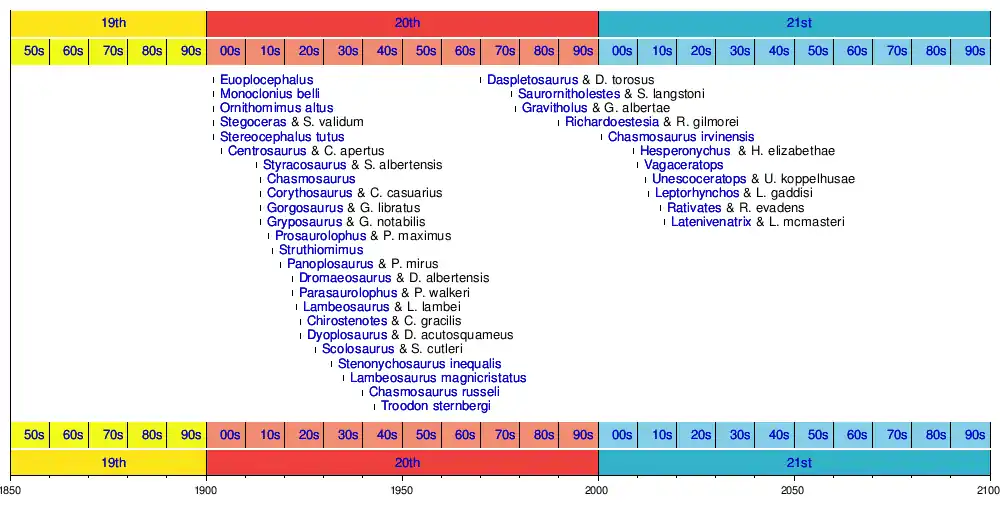
Timeline of new taxa
The following timeline displays valid taxa first discovered in the dinosaur. Some species may have been referred to other genera subsequent to their initial description.
See also
- List of dinosaur-bearing rock formations
Footnotes
- Lexicon of Canadian Geologic Units: Dinosaur Park Formation Archived 2013-02-21 at Archive.today
- Eberth, D.A.; Hamblin, A.P. (1993). "Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana". Canadian Journal of Earth Sciences. 30: 174–200. doi:10.1139/e93-016.
- Fowler, D. (2016). A new correlation of the Cretaceous formations of the Western Interior of the United States, I: Santonian-Maastrichtian formations and dinosaur biostratigraphy. Peer J Preprints.
- Eberth, D.A. 2005. The geology. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p.54-82. ISBN 0-253-34595-2.
- Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 277-291. ISBN 0-253-34595-2.
- Ryan and Evans (2005).
- Evans D.C.; Bavington R.; Campione N.E. (2009). "An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta". Canadian Journal of Earth Sciences. 46 (11): 791–800. Bibcode:2009CaJES..46..791E. doi:10.1139/E09-050.
- Arbour, V. M.; Burns, M. E.; Sissons, R. L. (2009). "A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus". Journal of Vertebrate Paleontology. 29 (4): 1117–1135. doi:10.1671/039.029.0405. S2CID 85665879.
- Penkalski, P. (2013). "A new ankylosaurid from the late Cretaceous Two Medicine Formation of Montana, USA". Acta Palaeontologica Polonica. doi:10.4202/app.2012.0125.
- Mallon, J. C., Evans, D. C., Ryan, M. J., & Anderson, J. S. (2012). Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology.
- Gardner, J.D. 2005. Lissamphibians. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 186-201. ISBN 0-253-34595-2.
- "Fossilworks: Gateway to the Paleobiology Database". fossilworks.org. Retrieved 23 April 2018.
- "Fossilworks: Gateway to the Paleobiology Database". fossilworks.org. Retrieved 23 April 2018.
- Currie, P.J. 2005. Theropods, including birds. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 367-397. ISBN 0-253-34595-2.
- Ryan, M.J., and Evans, D.C. 2005. Ornithischian dinosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 312-348. ISBN 0-253-34595-2.
- "Table 17.1," in Weishampel, et al. (2004). Page 365.
- Penkalski, Paul (2018). "Revised systematics of the armoured dinosaur Euoplocephalus and its allies". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 287 (3): 261–306. doi:10.1127/njgpa/2018/0717.
- Penkalski, Paul (2013). "A new ankylosaurid from the late Cretaceous Two Medicine Formation of Montana, USA". Acta Palaeontologica Polonica. doi:10.4202/app.2012.0125.
- "Table 23.1," in Weishampel, et al. (2004). Page 495.
- "Table 23.1," in Weishampel, et al. (2004). Page 496.
- Ryan, Michael J.; Evans, David C.; Currie, Phillip J.; Loewen, Mark A. (2014). "A New chasmosaurine from northern Laramidia expands frill disparity in ceratopsid dinosaurs". Naturwissenschaften. 101 (6): 505–512. doi:10.1007/s00114-014-1183-1. PMID 24859020. S2CID 13957187.
- Nicholas R. Longrich (2014). "The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography". Cretaceous Research. 51: 292–308. doi:10.1016/j.cretres.2014.06.011.
- Jordan C. Mallon, Christopher J. Ott, Peter L. Larson, Edward M. Iuliano and David C. Evans (2016). "Spiclypeus shipporum gen. et sp. nov., a Boldly Audacious New Chasmosaurine Ceratopsid (Dinosauria: Ornithischia) from the Judith River Formation (Upper Cretaceous: Campanian) of Montana, USA". PLOS ONE. 11 (5): e0154218. Bibcode:2016PLoSO..1154218M. doi:10.1371/journal.pone.0154218. PMC 4871577. PMID 27191389.CS1 maint: uses authors parameter (link)
- Farke, Andrew A.; Michael J. Ryan; Paul M. Barrett; Darren H. Tanke; Dennis R. Braman; Mark A. Loewen; Mark R. Graham (2011). "A new centrosaurine from the Late Cretaceous of Alberta, Canada, and the evolution of parietal ornamentation in horned dinosaurs" (PDF). Acta Palaeontologica Polonica. 56 (4): 691–702. doi:10.4202/app.2010.0121. S2CID 13717580.
- Michael J. Ryan; David C. Evans; Philip J. Currie; Caleb M. Brown; Don Brinkman (2012). "New leptoceratopsids from the Upper Cretaceous of Alberta, Canada". Cretaceous Research. 35: 69–80. doi:10.1016/j.cretres.2011.11.018.
- Scott D. Sampson; Mark A. Loewen; Andrew A. Farke; Eric M. Roberts; Catherine A. Forster; Joshua A. Smith; Alan L. Titus (2010). "New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endemism". PLOS ONE. 5 (9): e12292. Bibcode:2010PLoSO...512292S. doi:10.1371/journal.pone.0012292. PMC 2929175. PMID 20877459.
- "Abstract," Tanke and Brett-Surman (2001). Page 206.
- "Conclusions," Tanke and Brett-Surman (2001). Page 212.
- "Eggshell," Tanke and Brett-Surman (2001). Page 209.
- "Introduction," Tanke and Brett-Surman (2001). Page 208.
- "Discussion," Tanke and Brett-Surman (2001). Page 212.
- "Table 20.1," in Weishampel, et al. (2004). Page 441.
- "Table 20.1," in Weishampel, et al. (2004). Page 440.
- "Table 20.1," in Weishampel, et al. (2004). Page 442.
- "Table 21.1," in Weishampel, et al. (2004). Page 465.
- "Introduction," Jacobsen (2001). Page 59.
- "Discussion," Jacobsen (2001). Page 61.
- Longrich, N. R. (2014). "The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography". Cretaceous Research. 51: 292. doi:10.1016/j.cretres.2014.06.011.
- Longrich, N (2008). "A new, large ornithomimid from the Cretaceous Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains". Palaeontology. 51 (4): 983–997. doi:10.1111/j.1475-4983.2008.00791.x.
- McFeeters, B.; Ryan, M. J.; Schröder-Adams, C.; Currie, P. J. (2017). "First North American occurrences of Qiupalong (Theropoda: Ornithomimidae) and the palaeobiogeography of derived ornithomimids". FACETS. 2 (1): 355–373. doi:10.1139/facets-2016-0074.
- McFeeters, B.; et al. (2016). "A new ornithomimid theropod from the Dinosaur Park Formation of Alberta, Canada". Journal of Vertebrate Paleontology. 36 (6): e1221415. doi:10.1080/02724634.2016.1221415. S2CID 89242374.
- Longrich, N. R.; Barnes, K.; Clark, S.; Millar, L. (2013). "Caenagnathidae from the Upper Campanian Aguja Formation of West Texas, and a Revision of the Caenagnathinae". Bulletin of the Peabody Museum of Natural History. 54: 23–49. doi:10.3374/014.054.0102. S2CID 128444961.
- Funston, Gregory (2020-07-27). "Caenagnathids of the Dinosaur Park Formation (Campanian) of Alberta, Canada: anatomy, osteohistology, taxonomy, and evolution". Vertebrate Anatomy Morphology Palaeontology. 8: 105–153. doi:10.18435/vamp29362. ISSN 2292-1389.
- Sankey, Julia T.; Brinkman, Donald B.; Guenther, Merrilee; Currie, Philip J. (2002). "SMALL THEROPOD AND BIRD TEETH FROM THE LATE CRETACEOUS (LATE CAMPANIAN) JUDITH RIVER GROUP, ALBERTA". Journal of Paleontology. 76 (4): 751–763. doi:10.1666/0022-3360(2002)0762.0.CO;2. ISSN 0022-3360.
- Currie, Philip J.; Larson, Derek W. (2013-01-23). "Multivariate Analyses of Small Theropod Dinosaur Teeth and Implications for Paleoecological Turnover through Time". PLOS ONE. 8 (1): e54329. Bibcode:2013PLoSO...854329L. doi:10.1371/journal.pone.0054329. ISSN 1932-6203. PMC 3553132. PMID 23372708.
- "Abstract," Jacobsen (2001). Page 58.
- Currie, Philip J. (2003). "Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta". Acta Palaeontologica Polonica. 48 (2): 191–226.
- K.Gao and Brinkman, D.B. 2005. Choristoderes from the Park and its vicinity. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 221-234. ISBN 0-253-34595-2.
- Xiao-Chun Wu. 2005. Crocodylians. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 277-291. ISBN 0-253-34595-2.
- Caldwell, M.W. The squamates: origins, phylogeny, and paleoecology. In: Currie, P.J., and Koppelhus, E.B. (eds). 2005. ‘’Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 235-248. ISBN 0-253-34595-2.
- Sato, T., Eberth, D.A., Nicholls, E.L., and Manabe, M. 2005. Plesiosaurian remains from non-marine to paralic sediments. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 249-276. ISBN 0-253-34595-2.
- Godfrey, S.J., and Currie, P.J. 2005. Pterosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 292-311. ISBN 0-253-34595-2.
- David W. E. Hone; Michael B. Habib; François Therrien (2019). "Cryodrakon boreas gen. et sp. nov. a Late Cretaceous Canadian azhdarchid pterosaur". Journal of Vertebrate Paleontology. 39 (3): e1649681. doi:10.1080/02724634.2019.1649681. S2CID 203406859.
- Brinkman, D.B. 2005. Turtles: diversity, paleoecology, and distribution. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 202-220. ISBN 0-253-34595-2.
- Fox, R.C. 2005. Late Cretaceous mammals. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 417-435. ISBN 0-253-34595-2.
- Neuman, A.G., and Brinkman, D.B. 2005. Fishes of the fluvial beds. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 167-185. ISBN 0-253-34595-2.
- "Fossilworks: Gateway to the Paleobiology Database". fossilworks.org. Retrieved 23 April 2018.
- Hiroki Sato; Alison M. Murray; Oksana Vernygora; Philip J. Currie (2018). "A rare, articulated sturgeon (Chondrostei: Acipenseriformes) from the Upper Cretaceous of Dinosaur Provincial Park, Alberta, Canada". Journal of Vertebrate Paleontology. Online edition (4): (1)-(15). doi:10.1080/02724634.2018.1488137. S2CID 92574712.
- Johnston, P.A., and Hendy, A.J.W. 2005. Paleoecology of mollusks from the Upper Cretaceous Belly River Group. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 139-166. ISBN 0-253-34595-2.
- Koppelhus, E.B. 2005. Paleobotany. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 131-138. ISBN 0-253-34595-2.
- Braman, D.R., and Koppelhus, E.B. 2005. Campanian palynomorphs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 101-130. ISBN 0-253-34595-2.
References
- Arbour, V. M.; Burns, M. E.; Sissons, R. L. (2009). "A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus". Journal of Vertebrate Paleontology. 29 (4): 1117–1135. doi:10.1671/039.029.0405. S2CID 85665879.
- Braman, D.R., and Koppelhus, E.B. 2005. Campanian palynomorphs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 101-130.
- Brinkman, D.B. 2005. Turtles: diversity, paleoecology, and distribution. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 202-220.
- Caldwell, M.W. The squamates: origins, phylogeny, and paleoecology. In: Currie, P.J., and Koppelhus, E.B. (eds). 2005. ‘’Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 235-248.
- Currie, P.J. 2005. Theropods, including birds. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 367-397.
- Currie, P.J., and Koppelhus, E.B. (eds). 2005. Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 648 p.
- Eberth, D.A. 2005. The geology. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 54-82.
- Fox, R.C. 2005. Late Cretaceous mammals. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 417-435.
- K. Gao and Brinkman, D.B. 2005. Choristoderes from the Park and its vicinity. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 221-234.
- Gardner, J.D. 2005. Lissamphibians. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 186-201.
- Godfrey, S.J., and Currie, P.J. 2005. Pterosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 292-311.
- Johnston, P.A., and Hendy, A.J.W. 2005. Paleoecology of mollusks from the Upper Cretaceous Belly River Group. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 139-166.
- Koppelhus, E.B. 2005. Paleobotany. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 131-138.
- Lexicon of Canadian Geologic Units. "Dinosaur Park Formation". Archived from the original on 2013-02-21. Retrieved 2011-03-29.
- Neuman, A.G., and Brinkman, D.B. 2005. Fishes of the fluvial beds. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 167-185.
- Ryan, M.J., and Evans, D.C. 2005. Ornithischian dinosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 312-348.
- Sato, T., Eberth, D.A., Nicholls, E.L., and Manabe, M. 2005. Plesiosaurian remains from non-marine to paralic sediments. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 249-276.
- Tanke, D.H. and Brett-Surman, M.K. 2001. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206–218. In: Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Edited by D.H. Tanke and K. Carpenter. Indiana University Press: Bloomington. xviii + 577 pp.
- Xiao-Chun Wu. 2005. Crocodylians. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, 277-291