Macromolecular assembly
The term macromolecular assembly (MA) refers to massive chemical structures such as viruses and non-biologic nanoparticles, cellular organelles and membranes and ribosomes, etc. that are complex mixtures of polypeptide, polynucleotide, polysaccharide or other polymeric macromolecules. They are generally of more than one of these types, and the mixtures are defined spatially (i.e., with regard to their chemical shape), and with regard to their underlying chemical composition and structure. Macromolecules are found in living and nonliving things, and are composed of many hundreds or thousands of atoms held together by covalent bonds; they are often characterized by repeating units (i.e., they are polymers). Assemblies of these can likewise be biologic or non-biologic, though the MA term is more commonly applied in biology, and the term supramolecular assembly is more often applied in non-biologic contexts (e.g., in supramolecular chemistry and nanotechnology). MAs of macromolecules are held in their defined forms by non-covalent intermolecular interactions (rather than covalent bonds), and can be in either non-repeating structures (e.g., as in the ribosome (image) and cell membrane architectures), or in repeating linear, circular, spiral, or other patterns (e.g., as in actin filaments and the flagellar motor, image). The process by which MAs are formed has been termed molecular self-assembly, a term especially applied in non-biologic contexts. A wide variety of physical/biophysical, chemical/biochemical, and computational methods exist for the study of MA; given the scale (molecular dimensions) of MAs, efforts to elaborate their composition and structure and discern mechanisms underlying their functions are at the forefront of modern structure science.
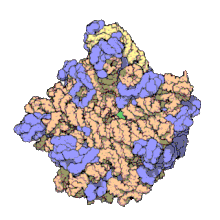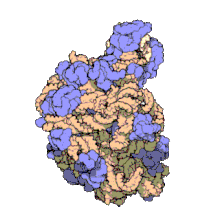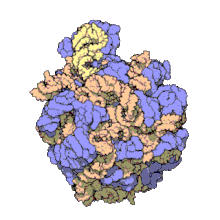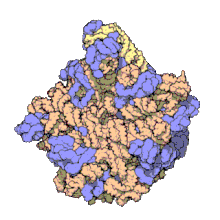
Biomolecular complex

A biomolecular complex, also called a biomacromolecular complex, is any biological complex made of more than one biopolymer (protein, RNA, DNA, [5] carbohydrate) or large non-polymeric biomolecules (lipid). The interactions between these biomolecules are non-covalent. [6] Examples:
- Protein complexes, some of which are multienzyme complexes: proteasome, DNA polymerase III holoenzyme, RNA polymerase II holoenzyme, symmetric viral capsids, chaperonin complex GroEL-GroES, photosystem I, ATP synthase, ferritin.
- RNA-protein complexes: ribosome, spliceosome, vault, SnRNP. Such complexes in cell nucleus are called ribonucleoproteins (RNPs).
- DNA-protein complexes: nucleosome.
- Protein-lipid complexes: lipoprotein.[7][8]
The biomacromolecular complexes are studied structurally by X-ray crystallography, NMR spectroscopy of proteins, cryo-electron microscopy and successive single particle analysis, and electron tomography. [9] The atomic structure models obtained by X-ray crystallography and biomolecular NMR spectroscopy can be docked into the much larger structures of biomolecular complexes obtained by lower resolution techniques like electron microscopy, electron tomography, and small-angle X-ray scattering. [10]
Complexes of macromolecules occur ubiquitously in nature, where they are involved in the construction of viruses and all living cells. In addition, they play fundamental roles in all basic life processes (protein translation, cell division, vesicle trafficking, intra- and inter-cellular exchange of material between compartments, etc.). In each of these roles, complex mixtures of become organized in specific structural and spatial ways. While the individual macromolecules are held together by a combination of covalent bonds and intramolecular non-covalent forces (i.e., associations between parts within each molecule, via charge-charge interactions, van der Waals forces, and dipole-dipole interactions such as hydrogen bonds), by definition MAs themselves are held together solely via the noncovalent forces, except now exerted between molecules (i.e., intermolecular interactions).
MA scales and examples
The images above give an indication of the compositions and scale (dimensions) associated with MAs, though these just begin to touch on the complexity of the structures; in principle, each living cell is composed of MAs, but is itself an MA as well. In the examples and other such complexes and assemblies, MAs are each often millions of daltons in molecular weight (megadaltons, i.e., millions of times the weight of a single, simple atom), though still having measurable component ratios (stoichiometries) at some level of precision. As alluded to in the image legends, when properly prepared, MAs or component subcomplexes of MAs can often be crystallized for study by protein crystallography and related methods, or studied by other physical methods (e.g., spectroscopy, microscopy).
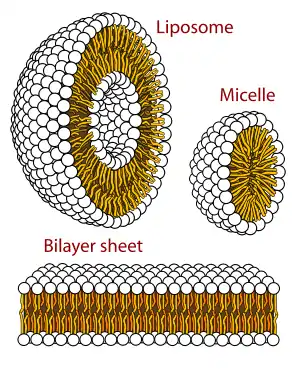
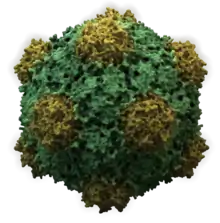
Virus structures were among the first studied MAs; other biologic examples include ribosomes (partial image above), proteasomes, and translation complexes (with protein and nucleic acid components), procaryotic and eukaryotic transcription complexes, and nuclear and other biological pores that allow material passage between cells and cellular compartments. Biomembranes are also generally considered MAs, though the requirement for structural and spatial definition is modified to accommodate the inherent molecular dynamics of membrane lipids, and of proteins within lipid bilayers.
Research into MAs
The study of MA structure and function is challenging, in particular because of their megadalton size, but also because of their complex compositions and varying dynamic natures. Most have had standard chemical and biochemical methods applied (methods of protein purification and centrifugation, chemical and electrochemical characterization, etc.). In addition, their methods of study include modern proteomic approaches, computational and atomic-resolution structural methods (e.g., X-ray crystallography), small-angle X-ray scattering (SAXS) and small-angle neutron scattering (SANS), force spectroscopy, and transmission electron microscopy and cryo-electron microscopy. Aaron Klug was recognized with the 1982 Nobel Prize in Chemistry for his work on structural elucidation using electron microscopy, in particular for protein-nucleic acid MAs including the tobacco mosaic virus (a structure containing a 6400 base ssRNA molecule and >2000 coat protein molecules). The crystallization and structure solution for the ribosome, MW ~ 2.5 MDa, an example of part of the protein synthetic 'machinery' of living cells, was object of the 2009 Nobel Prize in Chemistry awarded to Venkatraman Ramakrishnan, Thomas A. Steitz, and Ada E. Yonath.
Non-biologic counterparts
Finally, biology is not the sole domain of MAs. The fields of supramolecular chemistry and nanotechnology each have areas that have developed to elaborate and extend the principles first demonstrated in biologic MAs. Of particular interest in these areas has been elaborating the fundamental processes of molecular machines, and extending known machine designs to new types and processes.
See also
- Multi-state modeling of biomolecules
- Quaternary structure
- Multiprotein complex
- Organelle: the broadest definition of "organelle" includes not only membrane bound cellular structures, but also very large biomolecular complexes.
- Multi-state modeling of biomolecules
References
- Ban N, Nissen P, Hansen J, Moore P, Steitz T (2000). "The Complete Atomic Structure of the Large Ribosomal Subunit at 2.4 ångström Resolution". Science. 289 (5481): 905–20. Bibcode:2000Sci...289..905B. CiteSeerX 10.1.1.58.2271. doi:10.1126/science.289.5481.905. PMID 10937989.
- William McClure. "50S Ribosome Subunit". Archived from the original on 2005-11-24. Retrieved 2019-10-09.
- Osborne AR, Rapoport TA, van den Berg B (2005). "Protein translocation by the Sec61/SecY channel". Annual Review of Cell and Developmental Biology. 21: 529–50. doi:10.1146/annurev.cellbio.21.012704.133214. PMID 16212506.
- Legend, cover art, J. Bacteriol., October 2006.
- Kleinjung, Jens; Franca Fraternali (2005-07-01). "POPSCOMP: an automated interaction analysis of biomolecular complexes". Nucleic Acids Research. 33 (suppl 2): W342–W346. doi:10.1093/nar/gki369. ISSN 0305-1048. PMC 1160130. PMID 15980485. Retrieved 2013-11-14.
- Moore, Peter B. (2012). "How Should We Think About the Ribosome?". Annual Review of Biophysics. 41 (1): 1–19. doi:10.1146/annurev-biophys-050511-102314. PMID 22577819.
- Neuman, Nicole (January 2016). "The Complex Macromolecular Complex: Trends in Biochemical Sciences". Trends in Biochemical Sciences. 41 (1): 1–3. doi:10.1016/j.tibs.2015.11.006. PMID 26699226. Retrieved 2018-07-11.
- Dutta, Shuchismita; Berman, Helen M. (2005-03-01). "Large Macromolecular Complexes in the Protein Data Bank: A Status Report". Structure. 13 (3): 381–388. doi:10.1016/j.str.2005.01.008. ISSN 0969-2126. PMID 15766539.
- Russell, Robert B; Frank Alber; Patrick Aloy; Fred P Davis; Dmitry Korkin; Matthieu Pichaud; Maya Topf; Andrej Sali (June 2004). "A structural perspective on protein–protein interactions". Current Opinion in Structural Biology. 14 (3): 313–324. doi:10.1016/j.sbi.2004.04.006. ISSN 0959-440X. PMID 15193311.
- van Dijk, Aalt D. J.; Rolf Boelens; Alexandre M. J. J. Bonvin (2005). "Data-driven docking for the study of biomolecular complexes". FEBS Journal. 272 (2): 293–312. doi:10.1111/j.1742-4658.2004.04473.x. hdl:1874/336958. ISSN 1742-4658. PMID 15654870.
- "Structure of Fluid Lipid Bilayers". Blanco.biomol.uci.edu. 2009-11-10. Retrieved 2019-10-09.
- Experimental system, dioleoylphosphatidylcholine bilayers. The hydrophobic hydrocarbon region of the lipid is ~30 Å (3.0 nm) as determined by a combination of neutron and X-ray scattering methods; likewise, the polar/interface region (glyceryl, phosphate, and headgroup moieties, with their combined hydration) is ~15 Å (1.5 nm) on each side, for a total thickness about equal to the hydrocarbon region. See S.H. White references, preceding and following.
- Wiener MC & White SH (1992). "Structure of a fluid dioleoylphosphatidylcholine bilayer determined by joint refinement of x-ray and neutron diffraction data. III. Complete structure". Biophys. J. 61 (2): 434–447. Bibcode:1992BpJ....61..434W. doi:10.1016/S0006-3495(92)81849-0. PMC 1260259. PMID 1547331.
- Hydrocarbon dimensions vary with temperature, mechanical stress, PL structure and coformulants, etc. by single- to low double-digit percentages of these values.
Further reading
General reviews
- Williamson, J.R. (2008). "Cooperativity in macromolecular assembly". Nature Chemical Biology. 4 (8): 458–465. doi:10.1038/nchembio.102. PMID 18641626.
- Perrakis A, Musacchio A, Cusack S, Petosa C. Investigating a macromolecular complex: the toolkit of methods. J Struct Biol. 2011 Aug;175(2):106-12. doi: 10.1016/j.jsb.2011.05.014. Epub 2011 May 18. Review. PubMed PMID: 21620973.
- Dafforn TR. So how do you know you have a macromolecular complex? Acta Crystallogr D Biol Crystallogr. 2007 Jan;63(Pt 1):17-25. Epub 2006 Dec 13. Review. PubMed PMID: 17164522; PubMed Central PMCID: PMC2483502.
- Wohlgemuth I, Lenz C, Urlaub H. Studying macromolecular complex stoichiometries by peptide-based mass spectrometry. Proteomics. 2015 Mar;15(5-6):862-79. doi: 10.1002/pmic.201400466. Epub 2015 Feb 6. Review. PubMed PMID: 25546807; PubMed Central PMCID: PMC5024058.
- Sinha C, Arora K, Moon CS, Yarlagadda S, Woodrooffe K, Naren AP. Förster resonance energy transfer—An approach to visualize the spatiotemporal regulation of macromolecular complex formation and compartmentalized cell signaling. Biochim Biophys Acta. 2014 Oct;1840(10):3067-72. doi: 10.1016/j.bbagen.2014.07.015. Epub 2014 Jul 30. Review. PubMed PMID: 25086255; PubMed Central PMCID: PMC4151567.
- Berg, J. Tymoczko, J. and Stryer, L., Biochemistry. (W. H. Freeman and Company, 2002), ISBN 0-7167-4955-6
- Cox, M. and Nelson, D. L., Lehninger Principles of Biochemistry. (Palgrave Macmillan, 2004), ISBN 0-7167-4339-6
Reviews on particular MAs
- Valle M. Almost lost in translation. Cryo-EM of a dynamic macromolecular complex: the ribosome. Eur Biophys J. 2011 May;40(5):589-97. doi: 10.1007/s00249-011-0683-6. Epub 2011 Feb 19. Review. PubMed PMID: 21336521.
- Monie TP. The Canonical Inflammasome: A Macromolecular Complex Driving Inflammation. Subcell Biochem. 2017;83:43-73. doi: 10.1007/978-3-319-46503-6_2. Review. PubMed PMID: 28271472.
- Perino A, Ghigo A, Damilano F, Hirsch E. Identification of the macromolecular complex responsible for PI3Kgamma-dependent regulation of cAMP levels. Biochem Soc Trans. 2006 Aug;34(Pt 4):502-3. Review. PubMed PMID: 16856844.
Primary sources
- Lasker, K.; Förster, F.; Walzthoeni, T.; Villa, E.; Unverdorben, P.; Beck, F.; Aebersold, R.; Sali, A.; Baumeister, W. (2012). "Molecular architecture of the 26S proteasome holocomplex determined by an integrative approach". Proc Natl Acad Sci USA. 109 (5): 1380–7. Bibcode:2012PNAS..109.1380L. doi:10.1073/pnas.1120559109. PMC 3277140. PMID 22307589.
- Russel, D.; Lasker, K.; Webb, B.; Velázquez-Muriel, J.; Tjioe, E.; Schneidman-Duhovny, D.; Peterson, B.; Sali, A. (2012). "Putting the pieces together: integrative modeling platform software for structure determination of macromolecular assemblies". PLOS Biol. 10 (1): e1001244. doi:10.1371/journal.pbio.1001244. PMC 3260315. PMID 22272186.
- Barhoum S, Palit S, Yethiraj A. Diffusion NMR studies of macromolecular complex formation, crowding and confinement in soft materials. Prog Nucl Magn Reson Spectrosc. 2016 May;94-95:1-10. doi: 10.1016/j.pnmrs.2016.01.004. Epub 2016 Feb 4. Review. PubMed PMID: 27247282.
Other sources
- Nobel Prizes in Chemistry (2012), The Nobel Prize in Chemistry 2009, Venkatraman Ramakrishnan, Thomas A. Steitz, Ada E. Yonath, The Nobel Prize in Chemistry 2009, accessed 13 June 2011.
- Nobel Prizes in Chemistry (2012), The Nobel Prize in Chemistry 1982, Aaron Klug, The Nobel Prize in Chemistry 1982, accessed 13 June 2011.
External links
- Beck Group (2019), Structure and function of large macromolecular assemblies (Beck group home page), Beck Group - Structure and function of large molecular assemblies - EMBL, accessed 13 June 2011.
- DMA Group (2019), Dynamics of macromolecular assembly (DMA Group home page), Dynamics of Macromolecular Assembly Section | National Institute of Biomedical Imaging and Bioengineering, accessed 13 June 2011.