Insulin-degrading enzyme
Insulin-degrading enzyme, also known as IDE, is an enzyme.[4]
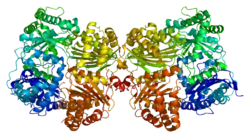


Known alternatively as insulysin or insulin protease, IDE is a large zinc-binding protease of the M16 metalloprotease family known to cleave multiple short polypeptides that vary considerably in sequence. Other members of this family include the mitochondrial processing peptidase[5] and presequence protease.[6]
Structure
Gene
The gene IDE encodes protein Insulin-degrading enzyme. The human gene IDE has 28 exons and is located at chromosome band 10q23-q25.[4]
Protein
Due to alternative splicing, The human protein Insulin-degrading Enzyme has two isoforms. Isoform1 is ~118 kDa in size and composed of 1019 amino acids while the isoform 2 is ~54.2 kDa [7] size and composed of 464 amino acids (missing 1-555 amino acids). The calculated theoretical pI of this protein isoform is 6.26.[8] Structural studies of IDE by Shen et al.[9] have provided insight into the functional mechanisms of the protease. Reminiscent of the previously determined structure of the bacterial protease pitrilysin, the IDE crystal structure reveals defined N and C terminal units that form a proteolytic chamber containing the zinc-binding active site. In addition, it appears that IDE can exist in two conformations: an open conformation, in which substrates can access the active site, and a closed state, in which the active site is contained within the chamber formed by the two concave domains. Targeted mutations that favor the open conformation result in a 40-fold increase in catalytic activity. Based upon this observation, it has been proposed that a possible therapeutic approach to Alzheimer’s might involve shifting the conformational preference of IDE to the open state, and thus increasing Aβ degradation, preventing aggregation, and, ideally, preventing the neuronal loss that leads to disease symptoms.[9]
Function
IDE was first identified by its ability to degrade the B chain of the hormone insulin. This activity was observed over sixty years ago,[10] though the enzyme specifically responsible for B chain cleavage was identified more recently.[11] This discovery revealed considerable amino acid sequence similarity between IDE and the previously characterized bacterial protease pitrilysin, suggesting a common proteolytic mechanism. IDE, which migrates at 110 kDa during gel electrophoresis under denaturing conditions, has since been shown to have additional substrates, including the signaling peptides glucagon, TGF alpha, and β-endorphin.[12]
Clinical Significance
Alzheimer's disease
Considerable interest in IDE has been stimulated due to the discovery that IDE can degrade amyloid beta (Aβ), a peptide implicated in the pathogenesis of Alzheimer's disease.[13] The underlying cause or causes of the disease are unclear, though the primary neuropathology observed is the formation of amyloid plaques and neurofibrillary tangles. One hypothesized mechanism of disease, called the amyloid hypothesis, suggests that the causative agent is the hydrophobic peptide Aβ, which forms quaternary structures that, by an unclear mechanism, cause neuronal death. Aβ is a byproduct generated as the result of proteolytic processing of the amyloid precursor protein (APP) by proteases referred to as the β and γ secretases. The physiological role of this processing is unclear, though it may play a role in nervous system development.[14]
Numerous in vitro and in vivo studies have shown correlations between IDE, Aβ degradation, and Alzheimer’s disease. Mice engineered to lack both alleles of the IDE gene exhibit a 50% decrease in Aβ degradation, resulting in cerebral accumulation of Aβ.[15] Studies of genetically inherited forms of Alzheimer’s show reduction in both IDE expression[16] and catalytic activity[17] among affected individuals. Despite the evident role of IDE in disease, relatively little is known about its physiological functions. These may be diverse, as IDE has been localized to several locations, including the cytosol, peroxisomes, endosomes, proteasome complexes,[18] and the surface of cerebrovascular endothelial cells.[19] Based upon the aforementioned observation in protein structure, it has been proposed that a possible therapeutic approach to Alzheimer’s might involve shifting the conformational preference of IDE to the open state, and thus increasing Aβ degradation, preventing aggregation, and, ideally, preventing the neuronal loss that leads to disease symptoms.
Regulation of extracellular amyloid β-protein
Reports of IDE localized to the cytosol and peroxisomes[20] have raised concerns regarding how the protease could degrade endogenous Aβ. Several studies have detected insulin-degrading activity in the conditioned media of cultured cells,[21][22] suggesting the permeability of the cell membrane and thus possible release of IDE from leaky cells. Qiu and colleagues revealed the presence of IDE in the extracellular media using antibodies to the enzyme. They also quantified levels of Aβ-degrading activity[23] using elution from column chromatography. Correlating the presence of IDE and Aβ-degrading activity in the conditioning medium confirmed that leaky membranes are responsible for extracellular IDE activity. However, other reports have indicated that it is released via exosomes.[24]
Potential role in the oligomerization of Aβ
Recent studies have observed that the oligomerization of synthetic Aβ was completely inhibited by the competitive IDE substrate, insulin.[23] These findings suggest that IDE activity is capable of joining of several Aβ fragments together. Qui et al. hypothesized that the Aβ fragments generated by IDE can either enhance oligomerization of the Aβ peptide or can oligomerize themselves. It is also entirely possible that IDE could mediate the degradation and oligomerization of Aβ by independent actions that have yet to be investigated.
Mechanism
The mechanism of the IDE enzyme remains poorly understood. The first step of one proposed mechanism[25] includes a zinc-bound hydroxide group performing a nucleophilic attack on a carbon substrate that materializes into the intermediate INT1. In this species, we can note that the zinc-bound hydroxide is completely transferred on the carbonyl carbon of substrate as a consequence of the Zn2+−OH bond breaking. In TS2, the Glu111 residue rotates to assume the right disposition to form two hydrogen bonds with the amide nitrogen and the −OH group linked to the carbon atom of substrate, thus behaving as hydrogen donor and acceptor, simultaneously. The formation of the second cited bond favors the re-establishment of the Zn2+−OH bond broken previously at the INT1 level. The nucleophilic addition and the protonation of peptide amide nitrogen is a very fast process that is believed to occur as a single step in the catalytic process. The final species on the path is the product PROD.[25] As a consequence of transfer of the proton of Glu111 onto the amide nitrogen of substrate that occurred in TS3, the peptide N—C bond is broken.
A look at the whole reaction path indicates that the rate-determining step in this process is the nucleophilic addition. After this point, the catalytic event should proceed without particular obstacles.[26][27]
Model organisms
| Characteristic | Phenotype |
|---|---|
| Homozygote viability | Normal |
| Fertility | Normal |
| General Observations | Abnormal |
| Body weight | Normal |
| Anxiety | Normal |
| Neurological assessment | Normal |
| Grip strength | Normal |
| Hot plate | Normal |
| Dysmorphology | Normal[28] |
| Indirect calorimetry | Normal |
| Glucose tolerance test | Normal |
| Auditory brainstem response | Normal |
| DEXA | Normal |
| Radiography | Normal |
| Body temperature | Normal |
| Eye morphology | Normal |
| Clinical chemistry | Normal |
| Haematology | Normal |
| Peripheral blood lymphocytes | Abnormal[29] |
| Micronucleus test | Normal |
| Heart weight | Normal |
| Salmonella infection | Normal[30] |
| Citrobacter infection | Normal[31] |
| All tests and analysis from[32][33] |
Model organisms have been used in the study of IDE function. A conditional knockout mouse line, called Idetm1a(EUCOMM)Wtsi[34][35] was generated as part of the International Knockout Mouse Consortium program — a high-throughput mutagenesis project to generate and distribute animal models of disease to interested scientists.[36][37][38]
Male and female animals underwent a standardized phenotypic screen to determine the effects of deletion.[32][39] Twenty three tests were carried out on mutant mice and two significant abnormalities were observed. Homozygous mutant animals displayed abnormal drinking behavior, and males also had an increased NK cell number.[32]
References
- GRCh38: Ensembl release 89: ENSG00000119912 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: IDE insulin-degrading enzyme".
- Aleshin AE, Gramatikova S, Hura GL, Bobkov A, Strongin AY, Stec B, et al. (November 2009). "Crystal and solution structures of a prokaryotic M16B peptidase: an open and shut case". Structure. 17 (11): 1465–75. doi:10.1016/j.str.2009.09.009. PMC 3615642. PMID 19913481.
- King JV, Liang WG, Scherpelz KP, Schilling AB, Meredith SC, Tang WJ (July 2014). "Molecular basis of substrate recognition and degradation by human presequence protease". Structure. 22 (7): 996–1007. doi:10.1016/j.str.2014.05.003. PMC 4128088. PMID 24931469.
- "IDE - Insulin-degrading enzyme - Homo sapiens (Human) - IDE gene & protein".
- "Uniprot: P14735 - IDE_HUMAN".
- Shen Y, Joachimiak A, Rosner MR, Tang WJ (October 2006). "Structures of human insulin-degrading enzyme reveal a new substrate recognition mechanism". Nature. 443 (7113): 870–4. Bibcode:2006Natur.443..870S. doi:10.1038/nature05143. PMC 3366509. PMID 17051221.
- Mirsky IA, Broh-Kahn RH (January 1949). "The inactivation of insulin by tissue extracts; the distribution and properties of insulin inactivating extracts". Archives of Biochemistry. 20 (1): 1–9. PMID 18104389.
- Affholter JA, Fried VA, Roth RA (December 1988). "Human insulin-degrading enzyme shares structural and functional homologies with E. coli protease III". Science. 242 (4884): 1415–8. Bibcode:1988Sci...242.1415A. doi:10.1126/science.3059494. PMID 3059494.
- Wang DS, Dickson DW, Malter JS (2006). "beta-Amyloid degradation and Alzheimer's disease". Journal of Biomedicine & Biotechnology. 2006 (3): 58406. doi:10.1155/JBB/2006/58406. PMC 1559921. PMID 17047308.
- Kurochkin IV, Goto S (May 1994). "Alzheimer's beta-amyloid peptide specifically interacts with and is degraded by insulin degrading enzyme". FEBS Letters. 345 (1): 33–7. doi:10.1016/0014-5793(94)00387-4. PMID 8194595. S2CID 43917847.
- Kerr ML, Small DH (April 2005). "Cytoplasmic domain of the beta-amyloid protein precursor of Alzheimer's disease: function, regulation of proteolysis, and implications for drug development". Journal of Neuroscience Research. 80 (2): 151–9. doi:10.1002/jnr.20408. PMID 15672415. S2CID 31985212.
- Farris W, Mansourian S, Chang Y, Lindsley L, Eckman EA, Frosch MP, et al. (April 2003). "Insulin-degrading enzyme regulates the levels of insulin, amyloid beta-protein, and the beta-amyloid precursor protein intracellular domain in vivo". Proceedings of the National Academy of Sciences of the United States of America. 100 (7): 4162–7. doi:10.1073/pnas.0230450100. PMC 153065. PMID 12634421.
- Cook DG, Leverenz JB, McMillan PJ, Kulstad JJ, Ericksen S, Roth RA, et al. (January 2003). "Reduced hippocampal insulin-degrading enzyme in late-onset Alzheimer's disease is associated with the apolipoprotein E-epsilon4 allele". The American Journal of Pathology. 162 (1): 313–9. doi:10.1016/s0002-9440(10)63822-9. PMC 1851126. PMID 12507914. Archived from the original on 2003-08-30. Retrieved 2008-02-10.
- Kim M, Hersh LB, Leissring MA, Ingelsson M, Matsui T, Farris W, et al. (March 2007). "Decreased catalytic activity of the insulin-degrading enzyme in chromosome 10-linked Alzheimer disease families". The Journal of Biological Chemistry. 282 (11): 7825–32. doi:10.1074/jbc.M609168200. PMID 17244626.
- Duckworth WC, Bennett RG, Hamel FG (October 1998). "Insulin degradation: progress and potential". Endocrine Reviews. 19 (5): 608–24. doi:10.1210/edrv.19.5.0349. PMID 9793760.
- Lynch JA, George AM, Eisenhauer PB, Conn K, Gao W, Carreras I, et al. (May 2006). "Insulin degrading enzyme is localized predominantly at the cell surface of polarized and unpolarized human cerebrovascular endothelial cell cultures". Journal of Neuroscience Research. 83 (7): 1262–70. doi:10.1002/jnr.20809. PMID 16511862. S2CID 23670388.
- Authier F, Bergeron JJ, Ou WJ, Rachubinski RA, Posner BI, Walton PA (April 1995). "Degradation of the cleaved leader peptide of thiolase by a peroxisomal proteinase". Proceedings of the National Academy of Sciences of the United States of America. 92 (9): 3859–63. Bibcode:1995PNAS...92.3859A. doi:10.1073/pnas.92.9.3859. PMC 42061. PMID 7731996.
- Roth RA, Mesirow ML, Cassell DJ, Yokono K, Baba S (March 1985). "Characterization of an insulin degrading enzyme from cultured human lymphocytes". Diabetes Research and Clinical Practice. 1 (1): 31–9. doi:10.1016/S0168-8227(85)80026-7. PMID 3915257.
- Semple JW, Lang Y, Speck ER, Delovitch TL (October 1992). "Processing and presentation of insulin. III. Insulin degrading enzyme: a neutral metalloendoproteinase that is non-homologous to classical endoproteinases mediates the processing of insulin epitopes for helper T cells". International Immunology. 4 (10): 1161–7. doi:10.1093/intimm/4.10.1161. PMID 1283335.
- Qiu WQ, Walsh DM, Ye Z, Vekrellis K, Zhang J, Podlisny MB, et al. (December 1998). "Insulin-degrading enzyme regulates extracellular levels of amyloid beta-protein by degradation". The Journal of Biological Chemistry. 273 (49): 32730–8. doi:10.1074/jbc.273.49.32730. PMID 9830016.
- Tamboli IY, Barth E, Christian L, Siepmann M, Kumar S, Singh S, et al. (November 2010). "Statins promote the degradation of extracellular amyloid {beta}-peptide by microglia via stimulation of exosome-associated insulin-degrading enzyme (IDE) secretion". The Journal of Biological Chemistry. 285 (48): 37405–14. doi:10.1074/jbc.M110.149468. PMC 2988346. PMID 20876579.
- Amata O, Marino T, Russo N, Toscano M (October 2009). "Human insulin-degrading enzyme working mechanism". Journal of the American Chemical Society. 131 (41): 14804–11. doi:10.1021/ja9037142. PMID 19785409.
- Leopoldini M, Russo N, Toscano M (August 2009). "Determination of the catalytic pathway of a manganese arginase enzyme through density functional investigation". Chemistry. 15 (32): 8026–36. doi:10.1002/chem.200802252. PMID 19288480.
- Hersh LB (November 2006). "The insulysin (insulin degrading enzyme) enigma". Cellular and Molecular Life Sciences. 63 (21): 2432–4. doi:10.1007/s00018-006-6238-9. PMID 16952049. S2CID 12536419.
- "Dysmorphology data for Ide". Wellcome Trust Sanger Institute.
- "Peripheral blood lymphocytes data for Ide". Wellcome Trust Sanger Institute.
- "Salmonella infection data for Ide". Wellcome Trust Sanger Institute.
- "Citrobacter infection data for Ide". Wellcome Trust Sanger Institute.
- Gerdin AK (2010). "The Sanger Mouse Genetics Programme: High throughput characterisation of knockout mice". Acta Ophthalmologica. 88: 925–7. doi:10.1111/j.1755-3768.2010.4142.x. S2CID 85911512.
- Mouse Resources Portal, Wellcome Trust Sanger Institute.
- "International Knockout Mouse Consortium".
- "Mouse Genome Informatics".
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, et al. (June 2011). "A conditional knockout resource for the genome-wide study of mouse gene function". Nature. 474 (7351): 337–42. doi:10.1038/nature10163. PMC 3572410. PMID 21677750.
- Dolgin E (June 2011). "Mouse library set to be knockout". Nature. 474 (7351): 262–3. doi:10.1038/474262a. PMID 21677718.
- Collins FS, Rossant J, Wurst W (January 2007). "A mouse for all reasons". Cell. 128 (1): 9–13. doi:10.1016/j.cell.2006.12.018. PMID 17218247. S2CID 18872015.
- van der Weyden L, White JK, Adams DJ, Logan DW (June 2011). "The mouse genetics toolkit: revealing function and mechanism". Genome Biology. 12 (6): 224. doi:10.1186/gb-2011-12-6-224. PMC 3218837. PMID 21722353.
External links
- The MEROPS online database for peptidases and their inhibitors: M16.002
- Insulin-Degrading+Enzyme at the US National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|