Nematothallus
Nematothallus is a form genus comprising cuticle-like fossils. Some of its constituents likely represent red algae,[3] whereas others resemble lichens.[2]
| Nematothallus | |
|---|---|
 | |
| Cuticle of Nematothallopsis showing reproductive apertures | |
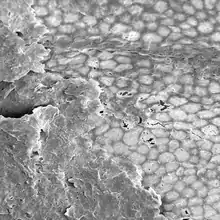 | |
| Cuticle of Nematothallopsis showing pseudocellular patterning | |
| Scientific classification | |
| Kingdom: | Plantae (?) |
| Phylum: | "Nematophyta" |
| Class: | †Nematophytina |
| Order: | †Nematophytales |
| Family: | †Nematothalaceae Strother (1993)[1] |
| Genus: | †Nematothallus Lang (1937) |
| Type species | |
| Nematothallus pseudovasculosa Lang (1937) | |
| Species | |
| |
History of research
Nematothallus was first described by Lang in 1937,[4] who envisioned it being an early thallose land plant with tubular features and sporophytes, covered by a cuticle which preserved impressions of the underlying cells.[5] He had found abundant disaggregated remains of all three features, none of which were connected to another, leaving his reconstruction of the phytodebris as parts of a single organism highly conjectural. Even so, it was picked up by Jonker (1973), who proposed that Nematothallus represented leaves of Prototaxites, which he interpreted as a red alga.[6]
Further work failed to draw together all aspects of the organism: Edwards (1982)[7] and Edwards and Rose (1984)[8] both provided thorough descriptions of the cuticular aspects of the plants, while Pratt et al. (1978)[9] and Niklas and Smocovitis (1983) focused on the anatomy of the tubes. Indeed, some workers suggested that the name Nematothallus should only apply to the tubes,[10][11] until Strother (1993) found more complete specimens, with tubes attached to the cuticle. He attempted to unite and formalise the genus, and extended it to include banded tubes,[1] which are instead referred to as nematoclasts (Graham & Gray 2001).[12]
It is possible that Nematothallus consisted of two layers of cuticle, although fossils giving this impression may in fact represent two layers which happened to overlap one another and become stuck. It is not readily established what the cuticle represents.[8] Lang (1945) had it as an epidermal layer, similar to the waxy cuticle of plants today, covering a parenchymatous layer. Alternatively, Edwards (1982) proposed that the inner tissue of Nematothallus comprised stringy tubes, with the cellular patterning produced by their ends.[7]
The genus was later formalised by Strother,[1] who discovered better preserved and more complete specimens in Pennsylvania, America – which appear to show tubes connected to the rims of cuticle.[11] Nematothallus is widespread from the late Silurian, but similar cuticle is reported from the Caradoc epoch (late Ordovician, 450 million years ago).[13] It is, however, difficult to distinguish Nematothallus cuticle from that of arthropods.[11]
Further work by Edwards and Rose has identified oval-shaped growths in places on the cuticles of a limited number of Nematothallus fragments, which develop into holes – whose purpose is unknown. Since they are not found in all Nematothallus individuals, it is unlikely that they were involved in gas exchange – perhaps they were involved in the release of spores? One fossil gives the impression, which may well be an illusion, of spores being trapped between two layers of cuticle.[8]
Habitat
Nematothallus was originally recovered from the Red Downtonian near the base of the Old Red Sandstone, between the clearly marine Ludlow deposits beneath it, and the unambiguously terrestrial Lower Old Red Sandstone.[4] Further samples came from elsewhere in the Lower Old Red Sandstone (St Maughams Group, lowermost Devonian), where they co-occur with plant spores.[8] Additional, older material occurs with Hostinella, Cooksonia and Steganotheca in the Ludlow (upper Ludfordian) Whitcliffe formation.[7]
Affinity
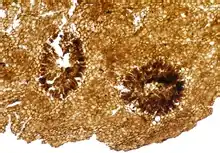
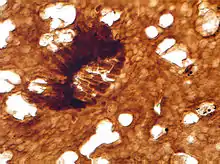
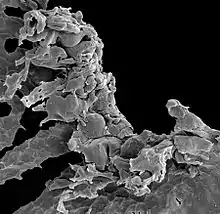
Some material assigned to Nematothallus closely resembles certain lichens, suggesting a lichenous habit – although as the lichens evolved at least six times independently, this does not fully establish the genetic affinity.[2] The biochemistry of the organism is not inconsistent with an algal affinity,[14] but Edwards (1982) considers it unlikely that algae would be preserved as coalified impressions.[7] However, Edwards does note that the surface patterning could have been produced in a similar fashion to surface layers in green algae – that is, by the ends of tightly packed filaments causing indentation on the surface layer. (Just because they were formed in the same way doesn't mean they were formed by green algae, though.)[7] The most likely affinity, for the aperture-bearing cuticles, is with the coralline red algae; the absence of biomineralization suggests a stem-group level affiliation.[3] Whereas Smith & Butterfield contend that this material is separate from the charcoalified lichen-like material, Edwards insists that all the material comes from the same lichen-like organism.[2]
Synonyms
N. taenia and N. lobata were both collapsed into a new form-species Laevitubulus laxus.[15]
See also
- Cosmochlaina, a closely related sister taxon.
- Spongiophyton, a similarly enigmatic organism with a similar cuticular patterning.[7] Some material described as Spongiophyton may in fact be Nematothallus.[16][17]
References
- Strother, P. K. (1993). "Clarification of the genus Nematothallus Lang". Journal of Paleontology. 67 (6): 1090–1094. doi:10.1017/S0022336000025476. JSTOR 1306131.
- Edwards, D.; Axe, L.; Honegger, R. (2013). "Contributions to the diversity in cryptogamic covers in the mid-Palaeozoic: Nematothallus revisited" (PDF). Botanical Journal of the Linnean Society. 173 (4): 505. doi:10.1111/boj.12119.
- Smith, M. R.; Butterfield, N. J. (2013). "A new view on Nematothallus: Coralline red algae from the Silurian of Gotland". Palaeontology. 56 (2): 345–357. doi:10.1111/j.1475-4983.2012.01203.x.
- Lang, W. H. (1937). "On the Plant-Remains from the Downtonian of England and Wales". Philosophical Transactions of the Royal Society B: Biological Sciences. 227 (544): 245–291. Bibcode:1937RSPTB.227..245L. doi:10.1098/rstb.1937.0004. JSTOR 92244.
- Lang, W. H. (1945). "Pachytheca and some anomalous early plants (Prototaxites, Nematothallus, Parka, Foerstia, Orvillea n.g.)". Botanical Journal of the Linnean Society. 52 (347): 535–552. doi:10.1111/j.1095-8339.1945.tb01351.x.
- Jonker, FP (1979). "Prototaxites in the lower Devonian". Palaeontographica Abteilung B. 171 (1–3): 39–56.
- Edwards, D. (1982). "Fragmentary non-vascular plant microfossils from the late Silurian of Wales". Botanical Journal of the Linnean Society. 84 (3): 223–256. doi:10.1111/j.1095-8339.1982.tb00536.x.
- Edwards, D.; Rose, V. (1984). "Cuticles of Nematothallus: a further enigma". Botanical Journal of the Linnean Society. 88 (1–2): 35–54. doi:10.1111/j.1095-8339.1984.tb01563.x.
- Pratt, L.; Phillips, T. L.; Dennison, J. M. (1978). "Evidence of non-vascular land plants from the early Silurian (Llandoverian) of Virginia, U.S.A". Review of Palaeobotany and Palynology. 25 (2): 121–149. doi:10.1016/0034-6667(78)90034-9.
- Gensel, P. G.; Johnson, N. G.; Strother, P. K. (1990). "Early Land Plant Debris (Hooker's "Waifs and Strays"?)". PALAIOS. 5 (6): 520–547. Bibcode:1990Palai...5..520G. doi:10.2307/3514860. JSTOR 3514860.
- Strother, P. K. (1988). "New Species of Nematothallus from the Silurian Bloomsburg Formation of Pennsylvania". Journal of Paleontology. 62 (6): 967–982. doi:10.1017/S0022336000030237. JSTOR 1305385.
- Graham, L.E.; Gray, J. (2001). The Origin, Morphology, and Ecophysiology of Early Embryophytes: Neontological and Paleontological Perspectives. Plants Invade the Land: Evolutionary and Environmental Perspectives. Columbia University Press. doi:10.7312/gens11160-009. ISBN 978-0-231-11161-4. Retrieved 2008-05-06.
- Edwards, D.; Kerp, H.; Hass, H. (1998). "Stomata in early land plants: an anatomical and ecophysiological approach". Journal of Experimental Botany. 49 (90001): 255. doi:10.1093/jexbot/49.suppl_1.255.
- Niklas, K. J. (1976). "Chemical Examinations of Some Non-Vascular Paleozoic Plants". Brittonia. 28 (1): 113–137. doi:10.2307/2805564. JSTOR 2805564.
- Burgess, N. D.; Edwards, D. (1991). "Classification of uppermost Ordovician to Lower Devonian tubular and filamentous macerals from the Anglo-Welsh Basin". Botanical Journal of the Linnean Society. 106: 41–66. doi:10.1111/j.1095-8339.1991.tb02282.x.
- "Spongiophyton z dolnego dewonu Polski" [On Spongiophyton from the Lower Devonian of Poland]. Acta Palaeobotanica (in Polish). 19 (1): 13–20. 1978.
- Filipiak, P.; Zatoń, M. (2010). "Plant and animal cuticle remains from the Lower Devonian of southern Poland and their palaeoenvironmental significance". Lethaia. 44 (4): 397–409. doi:10.1111/j.1502-3931.2010.00251.x.
- McGregor, DC; Narbonne, GM (1978). "Upper Silurian trilete spores and other microfossils from the Read Bay Formation, Cornwallis Island, Canadian Arctic". Canadian Journal of Earth Sciences. 15 (8): 1292–1303. Bibcode:1978CaJES..15.1292M. doi:10.1139/e78-136.