Serine C-palmitoyltransferase
In enzymology, a serine C-palmitoyltransferase (EC 2.3.1.50) is an enzyme that catalyzes the chemical reaction:[2][3]
- palmitoyl-CoA + L-serine CoA + 3-dehydro-D-sphinganine + CO2

| serine C-palmitoyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 2.3.1.50 | ||||||||
| CAS number | 62213-50-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
| Serine palmitoyltransferase | |||||||
|---|---|---|---|---|---|---|---|
 Crystallographic structure of serine palmitoyltransferase from S. paucimobilis. The cofactor PLP is visible in the center.[1] | |||||||
| Identifiers | |||||||
| Symbol | SPT1 | ||||||
| PDB | 2JG2 | ||||||
| UniProt | Q93UV0 | ||||||
| Other data | |||||||
| EC number | 2.3.1.50 | ||||||
| |||||||
| serine palmitoyltransferase, long chain base subunit 1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | SPTLC1 | ||||||
| Alt. symbols | HSN1 | ||||||
| NCBI gene | 10558 | ||||||
| HGNC | 11277 | ||||||
| OMIM | 605712 | ||||||
| RefSeq | NM_006415 | ||||||
| UniProt | O15269 | ||||||
| Other data | |||||||
| EC number | 2.3.1.50 | ||||||
| Locus | Chr. 9 q22.31 | ||||||
| |||||||
| serine palmitoyltransferase, long chain base subunit 2 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | SPTLC2 | ||||||
| NCBI gene | 9517 | ||||||
| HGNC | 11278 | ||||||
| OMIM | 605713 | ||||||
| RefSeq | NM_004863 | ||||||
| UniProt | O15270 | ||||||
| Other data | |||||||
| EC number | 2.3.1.50 | ||||||
| Locus | Chr. 14 q24.3 | ||||||
| |||||||
| serine palmitoyltransferase, long chain base subunit 3 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | SPTLC3 | ||||||
| Alt. symbols | C20orf38, SPTLC2L | ||||||
| NCBI gene | 55304 | ||||||
| HGNC | 16253 | ||||||
| OMIM | 611120 | ||||||
| RefSeq | NM_018327 | ||||||
| UniProt | Q9NUV7 | ||||||
| Other data | |||||||
| EC number | 2.3.1.50 | ||||||
| Locus | Chr. 20 p12.1 | ||||||
| |||||||
Thus, the two substrates of this enzyme are palmitoyl-CoA and L-serine, whereas its 3 products are CoA, 3-dehydro-D-sphinganine, and CO2.[4][5] This reaction is a key step in the biosynthesis of sphingosine which is a precursor of many other sphingolipids.[3]
This enzyme participates in sphingolipid metabolism. It employs one cofactor, pyridoxal phosphate.
Nomenclature
This enzyme belongs to the family of transferases, specifically those acyltransferases transferring groups other than aminoacyl groups. The systematic name of this enzyme class is palmitoyl-CoA:L-serine C-palmitoyltransferase (decarboxylating). Other names in common use include:
- serine palmitoyltransferase,
- SPT, 3-oxosphinganine synthetase, and
- acyl-CoA:serine C-2 acyltransferase decarboxylating.
Structure
Serine C-palmitoyltransferase is a member of the AOS (a-oxoamine synthase) family of PLP-dependent enzymes, which catalyse the condensation of amino acids and acyl-CoA thioester substrates.[6] The human enzyme is a heterodimer consisting of two monomeric subunits known as long chain base 1 and 2 (LCB1/2) encoded by separate genes.[1] The active site of LCB2 contains lysine and other key catalytic residues that are not present in LCB1, which does not participate in catalysis but is nevertheless required for the synthesis and stability of the enzyme.[7]
As of late 2007, two structures have been solved for this class of enzymes, with PDB accession codes 2JG2 and 2JGT.[1]
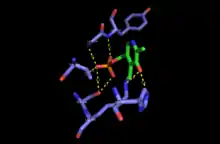
Mechanism
The PLP (pyridoxal 5′-phosphate)-dependent serine C-palmitoyltransferase carries out the first enzymatic step of de novo sphingolipid biosynthesis. The enzyme catalyses a Claisen-like condensation between L-serine and an acyl-CoA thioester (CoASH) substrate (typically C16-palmitoyl) or an acyl-ACP (acyl-carrier protein) thioester substrate, to form 3-ketodihydrosphingosine. Initially PLP cofactor is bound to the active-site lysine via a Schiff base to form the holo-form or internal aldimine of the enzyme. The amine group of L-serine then attacks and displaces the lysine bound to PLP, forming the external aldimine intermediate. Subsequently, deprotonation occurs at the Cα of serine, forming the quinonoid intermediate that attacks the incoming thioester substrate. Following decarboxylation and lysine attack, the product 3-keto-dihydrosphingosine is released and catalytically active PLP is reformed. This condensation reaction forms the sphingoid base or long-chain base found in all subsequent intermediate sphingolipids and complex sphingolipids in the organism.[3]
Isoforms
A variety of different serine C-palmitoyltransferase isoforms exist across different species. Unlike in eukaryotes, where the enzyme is heterodimeric and membrane bound, bacterial enzymes are homodimers and cytoplasmic. Studies of the isoform of the enzyme found in the Gram-negative bacterium S. paucimobilis were the first to elucidate the structure of the enzyme, revealing that PLP cofactor is held in place by several active site residues including Lys265 and His159.[8] Specifically, the S. paucimobilis isoform features an active-site arginine residue (Arg378) that plays a key role in stabilizing the carboxy moiety of the PLP-L-serine external aldimine intermediate. Similar arginine residues in enzyme homologues (Arg370, Arg390) play analogous roles.[9] Other homologues, such as in Sphingobacterium multivorum, feature the carboxy moiety bound to serine and methionine residues via water in place of arginine.[10] Certain enzyme homologues, such as in S. multivorum as well as B. stolpii, are found to be associated with the inner cell membrane, thus resembling the eukaryotic enzymes.[11] The B. stolpii homologue also features substrate inhibition by palmitoyl-CoA, a feature shared by the yeast and mammalian homologues.[12][13][14]
Clinical significance
HSAN1 (hereditary sensory and autonomic neuropathy type 1) is a genetic disorder caused by mutations in either one of SPTLC1 or SPTLC2, genes encoding the two heterodimeric subunits of the eukaryotic serine C-palmitoyltransferase enzyme.[15][16][17] These mutations have been shown to alter active site specificity, specifically by enhancing the ability of the enzyme to condense L-alanine with the palmitoyl-CoA substrate.[18] This is consistent with elevated levels of deoxysphingoid bases formed by the condensation of alanine with palmitoyl-CoA observed in HSAN1 patients.[19]
Species distribution
Serine C-palmitoyltransferase is expressed in a large number of species from bacteria to humans. The bacterial enzyme is a water-soluble homodimer[2] whereas in eukaryotes the enzyme is a heterodimer which is anchored to the endoplasmic reticulum.[3] Humans and other mammals express three paralogous subunits SPTLC1, SPTLC2, and SPTLC3. It was originally proposed that the functional human enzyme is a heterodimer between a SPTLC1 subunit and a second subunit which is either SPTLC2 or SPTLC3.[20] However more recent data suggest that the enzyme may exist as a larger complex, possibly an octamer, comprising all three subunits.[21]
References
- Yard BA, Carter LG, Johnson KA, Overton IM, Dorward M, Liu H, McMahon SA, Oke M, Puech D, Barton GJ, Naismith JH, Campopiano DJ (July 2007). "The structure of serine palmitoyltransferase; gateway to sphingolipid biosynthesis". Journal of Molecular Biology. 370 (5): 870–86. doi:10.1016/j.jmb.2007.04.086. PMID 17559874.
- Ikushiro H, Hayashi H, Kagamiyama H (April 2003). "Bacterial serine palmitoyltransferase: a water-soluble homodimeric prototype of the eukaryotic enzyme". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1647 (1–2): 116–20. doi:10.1016/S1570-9639(03)00074-8. PMID 12686119.
- Hanada K (June 2003). "Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1632 (1–3): 16–30. doi:10.1016/S1388-1981(03)00059-3. PMID 12782147.
- Brady RN, Di Mari SJ, Snell EE (January 1969). "Biosynthesis of sphingolipid bases. 3. Isolation and characterization of ketonic intermediates in the synthesis of sphingosine and dihydrosphingosine by cell-free extracts of Hansenula ciferri". The Journal of Biological Chemistry. 244 (2): 491–6. PMID 4388074.
- Stoffel W, LeKim D, Sticht G (May 1968). "Biosynthesis of dihydrosphingosine in vitro". Hoppe-Seyler's Zeitschrift für Physiologische Chemie. 349 (5): 664–70. doi:10.1515/bchm2.1968.349.1.664. PMID 4386961.
- Eliot AC, Kirsch JF (2004). "Pyridoxal phosphate enzymes: mechanistic, structural, and evolutionary considerations". Annual Review of Biochemistry. 73: 383–415. doi:10.1146/annurev.biochem.73.011303.074021. PMID 15189147.
- Han G, Gable K, Yan L, Natarajan M, Krishnamurthy J, Gupta SD, Borovitskaya A, Harmon JM, Dunn TM (December 2004). "The topology of the Lcb1p subunit of yeast serine palmitoyltransferase". The Journal of Biological Chemistry. 279 (51): 53707–16. doi:10.1074/jbc.M410014200. PMID 15485854.
- Shiraiwa Y, Ikushiro H, Hayashi H (June 2009). "Multifunctional role of His159in the catalytic reaction of serine palmitoyltransferase". The Journal of Biological Chemistry. 284 (23): 15487–95. doi:10.1074/jbc.M808916200. PMC 2786316. PMID 19346561.
- Lowther J, Charmier G, Raman MC, Ikushiro H, Hayashi H, Campopiano DJ (June 2011). "Role of a conserved arginine residue during catalysis in serine palmitoyltransferase" (PDF). FEBS Letters. 585 (12): 1729–34. doi:10.1016/j.febslet.2011.04.013. PMID 21514297. S2CID 25828713.
- Ikushiro H, Islam MM, Okamoto A, Hoseki J, Murakawa T, Fujii S, Miyahara I, Hayashi H (October 2009). "Structural insights into the enzymatic mechanism of serine palmitoyltransferase from Sphingobacterium multivorum". Journal of Biochemistry. 146 (4): 549–62. doi:10.1093/jb/mvp100. PMID 19564159.
- Ikushiro H, Islam MM, Tojo H, Hayashi H (August 2007). "Molecular characterization of membrane-associated soluble serine palmitoyltransferases from Sphingobacterium multivorum and Bdellovibrio stolpii". Journal of Bacteriology. 189 (15): 5749–61. doi:10.1128/JB.00194-07. PMC 1951810. PMID 17557831.
- Gable K, Slife H, Bacikova D, Monaghan E, Dunn TM (March 2000). "Tsc3p is an 80-amino acid protein associated with serine palmitoyltransferase and required for optimal enzyme activity". The Journal of Biological Chemistry. 275 (11): 7597–603. doi:10.1074/jbc.275.11.7597. PMID 10713067.
- Hanada K, Hara T, Nishijima M (March 2000). "Purification of the serine palmitoyltransferase complex responsible for sphingoid base synthesis by using affinity peptide chromatography techniques". The Journal of Biological Chemistry. 275 (12): 8409–15. doi:10.1074/jbc.275.12.8409. PMID 10722674.
- Lara PN, Moon J, Redman MW, Semrad TJ, Kelly K, Allen JW, Gitlitz BJ, Mack PC, Gandara DR (January 2015). "Relevance of platinum-sensitivity status in relapsed/refractory extensive-stage small-cell lung cancer in the modern era: a patient-level analysis of southwest oncology group trials". Journal of Thoracic Oncology. 10 (1): 110–5. doi:10.1097/JTO.0000000000000385. PMC 4320001. PMID 25490004.
- Bejaoui K, Wu C, Scheffler MD, Haan G, Ashby P, Wu L, de Jong P, Brown RH (March 2001). "SPTLC1 is mutated in hereditary sensory neuropathy, type 1". Nature Genetics. 27 (3): 261–2. doi:10.1038/85817. PMID 11242106. S2CID 34442339.
- Gable K, Han G, Monaghan E, Bacikova D, Natarajan M, Williams R, Dunn TM (March 2002). "Mutations in the yeast LCB1 and LCB2 genes, including those corresponding to the hereditary sensory neuropathy type I mutations, dominantly inactivate serine palmitoyltransferase". The Journal of Biological Chemistry. 277 (12): 10194–200. doi:10.1074/jbc.M107873200. PMID 11781309.
- Rotthier A, Auer-Grumbach M, Janssens K, Baets J, Penno A, Almeida-Souza L, Van Hoof K, Jacobs A, De Vriendt E, Schlotter-Weigel B, Löscher W, Vondráček P, Seeman P, De Jonghe P, Van Dijck P, Jordanova A, Hornemann T, Timmerman V (October 2010). "Mutations in the SPTLC2 subunit of serine palmitoyltransferase cause hereditary sensory and autonomic neuropathy type I". American Journal of Human Genetics. 87 (4): 513–22. doi:10.1016/j.ajhg.2010.09.010. PMC 2948807. PMID 20920666.
- Gable K, Gupta SD, Han G, Niranjanakumari S, Harmon JM, Dunn TM (July 2010). "A disease-causing mutation in the active site of serine palmitoyltransferase causes catalytic promiscuity". The Journal of Biological Chemistry. 285 (30): 22846–52. doi:10.1074/jbc.M110.122259. PMC 2906276. PMID 20504773.
- Penno A, Reilly MM, Houlden H, Laurá M, Rentsch K, Niederkofler V, Stoeckli ET, Nicholson G, Eichler F, Brown RH, von Eckardstein A, Hornemann T (April 2010). "Hereditary sensory neuropathy type 1 is caused by the accumulation of two neurotoxic sphingolipids". The Journal of Biological Chemistry. 285 (15): 11178–87. doi:10.1074/jbc.M109.092973. PMC 2856995. PMID 20097765.
- Hornemann T, Richard S, Rütti MF, Wei Y, von Eckardstein A (December 2006). "Cloning and initial characterization of a new subunit for mammalian serine-palmitoyltransferase". The Journal of Biological Chemistry. 281 (49): 37275–81. doi:10.1074/jbc.M608066200. PMID 17023427.
- Hornemann T, Wei Y, von Eckardstein A (July 2007). "Is the mammalian serine palmitoyltransferase a high-molecular-mass complex?". The Biochemical Journal. 405 (1): 157–64. doi:10.1042/BJ20070025. PMC 1925250. PMID 17331073.