Single cell sequencing
Single cell sequencing examines the sequence information from individual cells with optimized next-generation sequencing (NGS) technologies, providing a higher resolution of cellular differences and a better understanding of the function of an individual cell in the context of its microenvironment.[1] For example, in cancer, sequencing the DNA of individual cells can give information about mutations carried by small populations of cells. In development, sequencing the RNAs expressed by individual cells can give insight into the existence and behavior of different cell types.[2] In microbial systems, a population of the same species can appear to be genetically clonal, but single-cell sequencing of RNA or epigenetic modifications can reveal cell-to-cell variability that may help populations rapidly adapt to survive in changing environments.[3]
Background
A typical human cell consists of about 2 x 3.3 billion base pairs of DNA and 600 million bases of mRNA. Usually a mix of millions of cells are used in sequencing the DNA or RNA using traditional methods like Sanger sequencing or Illumina sequencing. By using deep sequencing of DNA and RNA from a single cell, cellular functions can be investigated extensively.[1] Like typical NGS experiments, the protocols of single cell sequencing generally contain the following steps: isolation of a single cell, nucleic acid extraction and amplification, sequencing library preparation, sequencing and bioinformatic data analysis. It is more challenging to perform single cell sequencing in comparison with sequencing from cells in bulk. The minimal amount of starting materials from a single cell make degradation, sample loss and contamination exert pronounced effects on quality of sequencing data. In addition, due to the picogram level of the amount of nucleic acids used,[4] heavy amplification is often needed during sample preparation of single cell sequencing, resulting in the uneven coverage, noise and inaccurate quantification of sequencing data.
Recent technical improvements make single cell sequencing a promising tool for approaching a set of seemingly inaccessible problems. For example, heterogeneous samples, rare cell types, cell lineage relationships, mosaicism of somatic tissues, analyses of microbes that cannot be cultured, and disease evolution can all be elucidated through single cell sequencing.[5] Single cell sequencing was selected as the method of the year 2013 by Nature Publishing Group.[6]
Single-cell genome (DNA) sequencing
Single-cell DNA genome sequencing involves isolating a single cell, amplifying the whole genome or region of interest, constructing sequencing libraries, and then applying next-generation DNA sequencing (ex. Illumina, Ion Torrent). In mammalian systems, single-cell DNA sequencing has been widely applied to study normal physiology and disease. Single-cell resolution can uncover the roles of genetic mosaicism or intra-tumor genetic heterogeneity in cancer development or treatment response.[7] In the context of microbiomes, a genome from a single unicellular organism is referred to as a single amplified genome (SAG). Advancements in single-cell DNA sequencing have enabled the collection of genomic data from uncultivated prokaryotic species present in complex microbiomes.[8] Although SAGs are characterized by low completeness and significant bias, recent computational advances have achieved the assembly of near-complete genomes from composite SAGs.[9] Data obtained from microorganisms might establish processes for culturing in the future.[10] Some of the genome assembly tools that can be used in single cell genome sequencing include: SPAdes, IDBA-UD, Cortex and HyDA.[11]
Methods
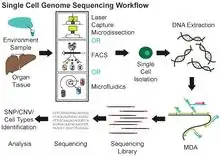
A list of more than 100 different single cell omics methods have been published.[12]
Multiple displacement amplification (MDA) is a widely used technique, enabling amplifying femtograms of DNA from bacterium to micrograms for the use of sequencing. Reagents required for MDA reactions include: random primers and DNA polymerase from bacteriophage phi29. In 30 degree isothermal reaction, DNA is amplified with included reagents. As the polymerases manufacture new strands, a strand displacement reaction takes place, synthesizing multiple copies from each template DNA. At the same time, the strands that were extended antecedently will be displaced. MDA products result in a length of about 12 kb and ranges up to around 100 kb, enabling its use in DNA sequencing.[10] In 2017, a major improvement to this technique, called WGA-X, was introduced by taking advantage of a thermostable mutant of the phi29 polymerase, leading to better genome recovery from individual cells, in particular those with high G+C content.[13] MDA has also been implemented in a microfluidic droplet-based system to achieve a highly parallelized single-cell whole genome amplification. By encapsulating single-cells in droplets for DNA capture and amplification, this method offers reduced bias and enhanced throughput compared to conventional MDA.[14]
Another common method is MALBAC.[15] This method begins with isothermal amplification as done in MDA, but the primers are flanked with a “common” sequence for downstream PCR amplification. As the preliminary amplicons are generated, the common sequence promotes self-ligation and the formation of “loops” to prevent further amplification. In contrast with MDA, the highly branched DNA network is not formed. Instead, in another temperature cycle, the loops are denatured, allowing the fragments to be amplified with PCR. MALBAC has also been implemented in a microfluidic device, but the amplification performance was not significantly improved by encapsulation in nanoliter droplets.[16]
Comparing MDA and MALBAC, MDA results in better genome coverage, but MALBAC provides more even coverage across the genome. MDA could be more effective for identifying SNPs, whereas MALBAC is preferred for detecting copy number variants. While performing MDA with a microfluidic device markedly reduces bias and contamination, the chemistry involved in MALBAC does not demonstrate the same potential for improved efficiency. The choice of method depends on the goal of the sequencing because each method presents different advantages.[7]
Limitations
MDA of individual cell genomes results in highly uneven genome coverage, i.e. relative overrepresentation and underrepresentation of various regions of the template, leading to loss of some sequences. There are two components to this process: a) stochastic over- and under-amplification of random regions; and b) systematic bias against high %GC regions. The stochastic component may be addressed by pooling single-cell MDA reactions from the same cell type, by employing fluorescent in situ hybridization (FISH) and/or post-sequencing confirmation.[10] The bias of MDA against high %GC regions can be addressed by using thermostable polymerases, such as in the process called WGA-X.[13]
Single-nucleotide polymorphisms (SNPs), which are a big part of genetic variation in the human genome, and copy number variation (CNV), pose problems in single cell sequencing, as well as the limited amount of DNA extracted from a single cell. Due to scant amounts of DNA, accurate analysis of DNA poses problems even after amplification since coverage is low and is susceptible to errors. With MDA, average genome coverage is less than 80% and SNPs that are not covered by sequencing reads will be opted out. In addition, MDA shows a high ratio of allele dropout, not detecting alleles from heterozygous samples. Various SNP algorithms are currently in use but none are specific to single cell sequencing. MDA with CNV also poses the problem of identifying false CNVs that conceal the real CNVs. To solve this, when patterns can be generated from false CNVs, algorithms can detect and eradicate this noise to produce true variants.[17]
Applications
Microbiomes are among the main targets of single cell genomics due to the difficulty of culturing the majority of microorganisms in most environments. Single-cell genomics is a powerful way to obtain microbial genome sequences without cultivation. This approach has been widely applied on marine, soil, subsurface, organismal, and other types of microbiomes in order to address a wide array of questions related to microbial ecology, evolution, public health and biotechnology potential.[18][19][20][21][22][23][24][25][26]
Cancer sequencing is also an emerging application of scDNAseq. Fresh or frozen tumors may be analyzed and categorized with respect to SCNAs, SNVs, and rearrangements quite well using whole genome DNAS approaches.[27] Cancer scDNAseq is particularly useful for examining the depth of complexity and compound mutations present in amplified therapeutic targets such as receptor tyrosine kinase genes (EGFR, PDGFRA etc.) where conventional population-level approaches of the bulk tumor are not able to resolve the co-occurrence patterns of these mutations within single cells of the tumor. Such overlap may provide redundancy of pathway activation and tumor cell resistance.
Single-cell DNA methylome sequencing
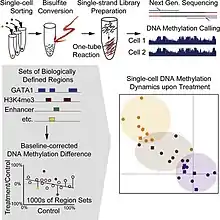
Single-cell DNA methylome sequencing quantifies DNA methylation. There are several known types of methylation that occur in nature, including 5-methylcytosine (5mC), 5-hydroymethylcytosine (5hmC), 6-methyladenine (6mA), and 4mC 4-methylcytosine (4mC). In eukaryotes, especially animals, 5mC is widespread along the genome and plays an important role in regulating gene expression by repressing transposable elements.[29] Sequencing 5mC in individual cells can reveal how epigenetic changes across genetically identical cells from a single tissue or population give rise to cells with different phenotypes.
Methods
Bisulfite sequencing has become the gold standard in detecting and sequencing 5mC in single cells.[30] Treatment of DNA with bisulfite converts cytosine residues to uracil, but leaves 5-methylcytosine residues unaffected. Therefore, DNA that has been treated with bisulfite retains only methylated cytosines. To obtain the methylome readout, the bisulfite-treated sequence is aligned to an unmodified genome. Whole genome bisulfite sequencing was achieved in single cells in 2014.[31] The method overcomes the loss of DNA associated with the typical procedure, where sequencing adapters are added prior to bisulfite fragmentation. Instead, the adapters are added after the DNA is treated and fragmented with bisulfite, allowing all fragments to be amplified by PCR.[32] Using deep sequencing, this method captures ~40% of the total CpGs in each cell. One way to improve the coverage of the method further would be to improve CpG capture efficiency by amplifying the DNA prior to bisulfite treatment.
Single-cell reduced representation bisulfite sequencing (scRRBS) is another method.[33] This method leverages the tendency of methylated cytosines to cluster at CpG islands (CGIs) to enrich for areas of the genome with a high CpG content. This reduces the cost of sequencing compared to whole genome bisulfite sequencing, but limits the coverage of this method. When RRBS is applied to bulk samples, majority of the CpG sites in gene promoters are detected, but site in gene promoters only account for 10% of CpG sites in the entire genome.[34] In single cells, 40% of the CpG sites from the bulk sample are detected. To increase coverage, this method can also be applied to a small pool of single cells. In a sample of 20 pooled single cells, 63% of the CpG sites from the bulk sample were detected. Pooling single cells is one strategy to increase methylome coverage, but at the cost of obscuring the heterogeneity in the population of cells.
Limitations
While bisulfite sequencing remains the most widely used approach for 5mC detection, the chemical treatment is harsh and fragments and degrades the DNA. This effect is exacerbated when moving from bulk samples to single cells. Other methods to detect DNA methylation include methylation-sensitive restriction enzymes. Restriction enzymes also enable the detection of other types of methylation, such as 6mA with DpnI.[35] Nanopore-based sequencing also offers a route for direct methylation sequencing without fragmentation or modification to the original DNA. Nanopore sequencing has been used to sequence the methylomes of bacteria, which are dominated by 6mA and 4mC (as opposed to 5mC in eukaryotes), but this technique has not yet been scaled down to single cells.[36]
Applications
Single-cell DNA methylation sequencing has been widely used to explore epigenetic differences in genetically similar cells. To validate these methods during their development, the single-cell methylome data of a mixed population were successfully classified by hierarchal clustering to identify distinct cell types.[33] Another application is studying single cells during the first few cell divisions in early development to understand how different cell types emerge from a single embryo.[37] Single-cell whole genome bisulfite sequencing has also been used to study rare but highly active cell types in cancer such as circulating tumor cells (CTCs).[38]

Single-cell assay for transposase-accessible chromatin with sequencing (scATAC-seq)
Single cell transposes-accessible chromatin sequencing maps chromatin accessibility across the genome. A transposase inserts sequencing adapters directly into open regions of chromatin, allowing those regions to be amplified and sequenced.[39]
Single-cell transcriptome sequencing (scRNA-seq)
Standard methods such as microarrays and bulk RNA-seq analysis analyze the expression of RNAs from large populations of cells. In mixed cell populations, these measurements may obscure critical differences between individual cells within these populations.[40][41]
Single-cell RNA sequencing (scRNA-seq) provides the expression profiles of individual cells and is considered the gold standard for defining cell states and phenotypes as of 2020.[42] Although it is not possible to obtain complete information on every RNA expressed by each cell, due to the small amount of material available, patterns of gene expression can be identified through gene clustering analyses.[43] This can uncover the existence of rare cell types within a cell population that may never have been seen before. For example, rare specialized cells in the lung called pulmonary ionocytes that express the Cystic Fibrosis Transmembrane Conductance Regulator were identified in 2018 by two groups performing scRNA-Seq on lung airway epithelia.[44][45]
Methods
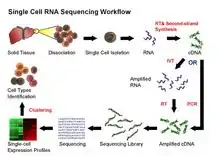
Current scRNA-seq protocols involve isolating single cells and their RNA, and then following the same steps as bulk RNA-seq: reverse transcription (RT), amplification, library generation and sequencing. Early methods separated individual cells into separate wells; more recent methods encapsulate individual cells in droplets in a microfluidic device, where the reverse transcription reaction takes place, converting RNAs to cDNAs. Each droplet carries a DNA "barcode" that uniquely labels the cDNAs derived from a single cell. Once reverse transcription is complete, the cDNAs from many cells can be mixed together for sequencing; transcripts from a particular cell are identified by the unique barcode.[46][47]
Challenges for scRNA-Seq include preserving the initial relative abundance of mRNA in a cell and identifying rare transcripts.[48] The reverse transcription step is critical as the efficiency of the RT reaction determines how much of the cell’s RNA population will be eventually analyzed by the sequencer. The processivity of reverse transcriptases and the priming strategies used may affect full-length cDNA production and the generation of libraries biased toward 3’ or 5' end of genes.
In the amplification step, either PCR or in vitro transcription (IVT) is currently used to amplify cDNA. One of the advantages of PCR-based methods is the ability to generate full-length cDNA. However, different PCR efficiency on particular sequences (for instance, GC content and snapback structure) may also be exponentially amplified, producing libraries with uneven coverage. On the other hand, while libraries generated by IVT can avoid PCR-induced sequence bias, specific sequences may be transcribed inefficiently, thus causing sequence drop-out or generating incomplete sequences.[1][40] Several scRNA-seq protocols have been published: Tang et al.,[49] STRT,[50] SMART-seq,[51] CEL-seq,[52] RAGE-seq,[53] Quartz-seq.[54] and C1-CAGE.[55] These protocols differ in terms of strategies for reverse transcription, cDNA synthesis and amplification, and the possibility to accommodate sequence-specific barcodes (i.e. UMIs) or the ability to process pooled samples.[56]
In 2017, two approaches were introduced to simultaneously measure single-cell mRNA and protein expression through oligonucleotide-labeled antibodies known as REAP-seq,[57] and CITE-seq.[58]
Limitations
Most RNA-Seq methods depend on poly(A) tail capture to enrich mRNA and deplete abundant and uninformative rRNA. Thus, they are often restricted to sequencing polyadenylated mRNA molecules. However, recent studies are now starting to appreciate the importance of non-poly(A) RNA, such as long-noncoding RNA and microRNAs in gene expression regulation. Small-seq is a single-cell method that captures small RNAs (<300 nucleotides) such as microRNAs, fragments of tRNAs and small nucleolar RNAs in mammalian cells.[59] This method uses a combination of “oligonucleotide masks” (that inhibit the capture of highly abundant 5.8S rRNA molecules) and size selection to exclude large RNA species such as other highly abundant rRNA molecules. To target larger non-poly(A) RNAs, such as long non-coding mRNA, histone mRNA, circular RNA, and enhancer RNA, size selection is not applicable for depleting the highly abundant ribosomal RNA molecules (18S and 28s rRNA).[60] Single-cell RamDA-Seq is a method that achieves this by performing reverse transcription with random priming (random displacement amplification) in the presence of “not so random” (NSR) primers specifically designed to avoid priming on rRNA molecule.[61] While this method successfully captures full-length total RNA transcripts for sequencing and detected a variety of non-poly(A) RNAs with high sensitivity, it has some limitations. The NSR primers were carefully designed according to rRNA sequences in the specific organism (mouse), and designing new primer sets for other species would take considerable effort.
Bacteria and other prokaryotes are currently not amenable to single-cell RNA-seq due to the lack of polyadenylated mRNA. Thus, the development of single-cell RNA-seq methods that do not depend on poly(A) tail capture will also be instrumental in enabling single-cell resolution microbiome studies. Bulk bacterial studies typically apply general rRNA depletion to overcome the lack of polyadenylated mRNA on bacteria, but at the single-cell level, the total RNA found in one cell is too small.[60] Lack of polyadenylated mRNA and scarcity of total RNA found in single bacteria cells are two important barriers limiting the deployment of scRNA-seq in bacteria.
Applications
scRNA-Seq is becoming widely used across biological disciplines including Developmental biology,[62] Neurology,[63] Oncology,[64][65][66] Immunology,[67][68] Cardiovascular research[69] and Infectious disease.[70][71]
Using machine learning methods, data from bulk RNA-Seq has been used to increase the signal/noise ratio in scRNA-Seq. Specifically, scientists have used gene expression profiles from pan-cancer datasets in order to build coexpression networks, and then have applied these on single cell gene expression profiles, obtaining a more robust method to detect the presence of mutations in individual cells using transcript levels.[72]
Some scRNA-seq methods have also been applied to single cell microorganisms. SMART-seq2 has been used to analyze single cell eukaryotic microbes, but since it relies on poly(A) tail capture, it has not been applied in prokaryotic cells.[73] Microfluidic approaches such as Drop-seq and the Fluidigm IFC-C1 devices have been used to sequence single malaria parasites or single yeast cells.[74][75] The single-cell yeast study sought to characterize the heterogeneous stress tolerance in isogenic yeast cells before and after the yeast are exposed to salt stress. Single-cell analysis of the several transcription factors by scRNA-seq revealed heterogeneity across the population. These results suggest that regulation varies among members of a population to increase the chances of survival for a fraction of the population.
The first single-cell transcriptome analysis in a prokaryotic species was accomplished using the terminator exonuclease enzyme to selectively degrade rRNA and rolling circle amplification (RCA) of mRNA.[76] In this method, the ends of single stranded DNA were ligated together to form a circle, and the resulting loop was then used as a template for linear RNA amplification. The final product library was then analyzed by microarray, with low bias and good coverage. However, RCA has not been tested with RNA-seq, which typically employs next-generation sequencing. Single-cell RNA-seq for bacteria would be highly useful for studying microbiomes. It would address issues encountered in conventional bulk metatranscriptomics approaches, such as failing to capture species present in low abundance, and failing to resolve heterogeneity among cell populations.
scRNA-Seq has provided considerable insight into the development of embryos and organisms, including the worm Caenorhabditis elegans,[77] and the regenerative planarian Schmidtea mediterranea[78][79] and axolotl Ambystoma mexicanum.[80][81] The first vertebrate animals to be mapped in this way were Zebrafish[82][83][84] and Xenopus laevis.[85] In each case multiple stages of the embryo were studied, allowing the entire process of development to be mapped on a cell-by-cell basis. Science recognized these advances as the 2018 Breakthrough of the Year.[86]
Considerations
Isolation of single cells
There are several ways to isolate individual cells prior to whole genome amplification and sequencing. Fluorescence-activated cell sorting (FACS) is a widely used approach. Individual cells can also be collected by micromanipulation, for example by serial dilution or by using a patch pipette or nanotube to harvest a single cell.[15][87] The advantages of micromanipulation are ease and low cost, but they are laborious and susceptible to misidentification of cell types under microscope. Laser-capture microdissection (LCM) can also be used for collecting single cells. Although LCM preserves the knowledge of the spatial location of a sampled cell within a tissue, it is hard to capture a whole single cell without also collecting the materials from neighboring cells.[40][88][89] High-throughput methods for single cell isolation also include microfluidics. Both FACS and microfluidics are accurate, automatic and capable of isolating unbiased samples. However, both methods require detaching cells from their microenvironments first, thereby causing perturbation to the transcriptional profiles in RNA expression analysis.[90][91]
scRNA-Seq
Generally speaking, for a typical bulk cell RNA-sequencing (RNA-seq) experiment, ten million reads are generated and a gene with higher than the threshold of 50 reads per kb per million reads (RPKM) is considered expressed. For a gene that is 1kb long, this corresponds to 500 reads and a minimum coefficient of variation (CV) of 4% under the assumption of the Poisson distribution. For a typical mammalian cell containing 200,000 mRNA, sequencing data from at least 50 single cells need to be pooled in order to achieve this minimum CV value. However, due to the efficiency of reverse transcription and other noise introduced in the experiments, more cells are required for accurate expression analyses and cell type identification.[40]
References
- Eberwine J, Sul JY, Bartfai T, Kim J (January 2014). "The promise of single-cell sequencing". Nature Methods. 11 (1): 25–7. doi:10.1038/nmeth.2769. PMID 24524134. S2CID 11575439.
- Pennisi E (April 2018). "Chronicling embryos, cell by cell, gene by gene". Science. 360 (6387): 367. Bibcode:2018Sci...360..367P. doi:10.1126/science.360.6387.367. PMID 29700246.
- Saliba AE, Westermann AJ, Gorski SA, Vogel J (August 2014). "Single-cell RNA-seq: advances and future challenges". Nucleic Acids Research. 42 (14): 8845–60. doi:10.1093/nar/gku555. PMC 4132710. PMID 25053837.
- Shintaku H, Nishikii H, Marshall LA, Kotera H, Santiago JG (February 2014). "On-chip separation and analysis of RNA and DNA from single cells". Analytical Chemistry. 86 (4): 1953–7. doi:10.1021/ac4040218. PMID 24499009.
- Nawy T (January 2014). "Single-cell sequencing". Nature Methods. 11 (1): 18. doi:10.1038/nmeth.2771. PMID 24524131. S2CID 5252333.
- "Method of the year 2013". Nature Methods. 11 (1): 1. January 2014. doi:10.1038/nmeth.2801. PMID 24524124.
- Gawad C, Koh W, Quake SR (March 2016). "Single-cell genome sequencing: current state of the science". Nature Reviews. Genetics. 17 (3): 175–88. doi:10.1038/nrg.2015.16. PMID 26806412. S2CID 4800650.
- Alneberg J, Karlsson CM, Divne AM, Bergin C, Homa F, Lindh MV, et al. (September 2018). "Genomes from uncultivated prokaryotes: a comparison of metagenome-assembled and single-amplified genomes". Microbiome. 6 (1): 173. doi:10.1186/s40168-018-0550-0. PMC 6162917. PMID 30266101.
- Kogawa M, Hosokawa M, Nishikawa Y, Mori K, Takeyama H (February 2018). "Obtaining high-quality draft genomes from uncultured microbes by cleaning and co-assembly of single-cell amplified genomes". Scientific Reports. 8 (1): 2059. Bibcode:2018NatSR...8.2059K. doi:10.1038/s41598-018-20384-3. PMC 5794965. PMID 29391438.
- "Lasken RS (October 2007). "Single-cell genomic sequencing using Multiple Displacement Amplification". Current Opinion in Microbiology. 10 (5): 510–6. doi:10.1016/j.mib.2007.08.005. PMID 17923430."
- Taghavi Z, Movahedi NS, Draghici S, Chitsaz H (October 2013). "Distilled single-cell genome sequencing and de novo assembly for sparse microbial communities". Bioinformatics. 29 (19): 2395–401. arXiv:1305.0062. Bibcode:2013arXiv1305.0062T. doi:10.1093/bioinformatics/btt420. PMC 3777112. PMID 23918251.
- "Single-Cell-Omics.v2.3.13 @albertvilella". Google Docs. Retrieved 2020-01-01.
- Stepanauskas R, Fergusson EA, Brown J, Poulton NJ, Tupper B, Labonté JM, et al. (July 2017). "Improved genome recovery and integrated cell-size analyses of individual uncultured microbial cells and viral particles". Nature Communications. 8 (1): 84. Bibcode:2017NatCo...8...84S. doi:10.1038/s41467-017-00128-z. PMC 5519541. PMID 28729688.
- Hosokawa M, Nishikawa Y, Kogawa M, Takeyama H (July 2017). "Massively parallel whole genome amplification for single-cell sequencing using droplet microfluidics". Scientific Reports. 7 (1): 5199. Bibcode:2017NatSR...7.5199H. doi:10.1038/s41598-017-05436-4. PMC 5507899. PMID 28701744.
- Zong C, Lu S, Chapman AR, Xie XS (December 2012). "Genome-wide detection of single-nucleotide and copy-number variations of a single human cell". Science. 338 (6114): 1622–6. Bibcode:2012Sci...338.1622Z. doi:10.1126/science.1229164. PMC 3600412. PMID 23258894.
- Yu Z, Lu S, Huang Y (October 2014). "Microfluidic whole genome amplification device for single cell sequencing". Analytical Chemistry. 86 (19): 9386–90. doi:10.1021/ac5032176. PMID 25233049.
- "Ning L, Liu G, Li G, Hou Y, Tong Y, He J (2014). "Current challenges in the bioinformatics of single cell genomics". Frontiers in Oncology. 4 (7): 7. doi:10.3389/fonc.2014.00007. PMC 3902584. PMID 24478987."
- Blainey PC, Quake SR (January 2014). "Dissecting genomic diversity, one cell at a time". Nature Methods. 11 (1): 19–21. doi:10.1038/nmeth.2783. hdl:1721.1/106574. PMC 3947563. PMID 24524132.
- Zhang K, Martiny AC, Reppas NB, Barry KW, Malek J, Chisholm SW, Church GM (June 2006). "Sequencing genomes from single cells by polymerase cloning". Nature Biotechnology. 24 (6): 680–6. doi:10.1038/nbt1214. PMID 16732271. S2CID 2994579.
- Yoon HS, Price DC, Stepanauskas R, Rajah VD, Sieracki ME, Wilson WH, et al. (May 2011). "Single-cell genomics reveals organismal interactions in uncultivated marine protists". Science. 332 (6030): 714–7. Bibcode:2011Sci...332..714Y. doi:10.1126/science.1203163. PMID 21551060. S2CID 34343205.
- Swan BK, Martinez-Garcia M, Preston CM, Sczyrba A, Woyke T, Lamy D, et al. (September 2011). "Potential for chemolithoautotrophy among ubiquitous bacteria lineages in the dark ocean". Science. 333 (6047): 1296–300. Bibcode:2011Sci...333.1296S. doi:10.1126/science.1203690. PMID 21885783. S2CID 206533092.
- Woyke T, Xie G, Copeland A, González JM, Han C, Kiss H, et al. (2009-04-23). "Assembling the marine metagenome, one cell at a time". PLOS ONE. 4 (4): e5299. Bibcode:2009PLoSO...4.5299W. doi:10.1371/journal.pone.0005299. PMC 2668756. PMID 19390573.
- Swan BK, Tupper B, Sczyrba A, Lauro FM, Martinez-Garcia M, González JM, et al. (July 2013). "Prevalent genome streamlining and latitudinal divergence of planktonic bacteria in the surface ocean". Proceedings of the National Academy of Sciences of the United States of America. 110 (28): 11463–8. Bibcode:2013PNAS..11011463S. doi:10.1073/pnas.1304246110. PMC 3710821. PMID 23801761.
- Rinke C, Schwientek P, Sczyrba A, Ivanova NN, Anderson IJ, Cheng JF, et al. (July 2013). "Insights into the phylogeny and coding potential of microbial dark matter" (PDF). Nature. 499 (7459): 431–7. Bibcode:2013Natur.499..431R. doi:10.1038/nature12352. PMID 23851394. S2CID 4394530.
- Kashtan N, Roggensack SE, Rodrigue S, Thompson JW, Biller SJ, Coe A, et al. (April 2014). "Single-cell genomics reveals hundreds of coexisting subpopulations in wild Prochlorococcus". Science. 344 (6182): 416–20. Bibcode:2014Sci...344..416K. doi:10.1126/science.1248575. hdl:1721.1/92763. PMID 24763590. S2CID 13659345.
- Pachiadaki MG, Sintes E, Bergauer K, Brown JM, Record NR, Swan BK, et al. (November 2017). "Major role of nitrite-oxidizing bacteria in dark ocean carbon fixation". Science. 358 (6366): 1046–1051. Bibcode:2017Sci...358.1046P. doi:10.1126/science.aan8260. PMID 29170234.
- Francis JM, Zhang CZ, Maire CL, Jung J, Manzo VE, Adalsteinsson VA, et al. (August 2014). "EGFR variant heterogeneity in glioblastoma resolved through single-nucleus sequencing". Cancer Discovery. 4 (8): 956–71. doi:10.1158/2159-8290.CD-13-0879. PMC 4125473. PMID 24893890.
- Farlik M, Sheffield NC, Nuzzo A, Datlinger P, Schönegger A, Klughammer J, Bock C (March 2015). "Single-cell DNA methylome sequencing and bioinformatic inference of epigenomic cell-state dynamics". Cell Reports. 10 (8): 1386–97. doi:10.1016/j.celrep.2015.02.001. PMC 4542311. PMID 25732828.
- Zemach A, McDaniel IE, Silva P, Zilberman D (May 2010). "Genome-wide evolutionary analysis of eukaryotic DNA methylation". Science. 328 (5980): 916–9. Bibcode:2010Sci...328..916Z. doi:10.1126/science.1186366. PMID 20395474. S2CID 206525166.
- Crary-Dooley FK, Tam ME, Dunaway KW, Hertz-Picciotto I, Schmidt RJ, LaSalle JM (March 2017). "A comparison of existing global DNA methylation assays to low-coverage whole-genome bisulfite sequencing for epidemiological studies". Epigenetics. 12 (3): 206–214. doi:10.1080/15592294.2016.1276680. PMC 5406214. PMID 28055307.
- Smallwood SA, Lee HJ, Angermueller C, Krueger F, Saadeh H, Peat J, et al. (August 2014). "Single-cell genome-wide bisulfite sequencing for assessing epigenetic heterogeneity". Nature Methods. 11 (8): 817–820. doi:10.1038/nmeth.3035. PMC 4117646. PMID 25042786.
- Miura F, Enomoto Y, Dairiki R, Ito T (September 2012). "Amplification-free whole-genome bisulfite sequencing by post-bisulfite adaptor tagging". Nucleic Acids Research. 40 (17): e136. doi:10.1093/nar/gks454. PMC 3458524. PMID 22649061.
- Guo H, Zhu P, Wu X, Li X, Wen L, Tang F (December 2013). "Single-cell methylome landscapes of mouse embryonic stem cells and early embryos analyzed using reduced representation bisulfite sequencing". Genome Research. 23 (12): 2126–35. doi:10.1101/gr.161679.113. PMC 3847781. PMID 24179143.
- Gu H, Smith ZD, Bock C, Boyle P, Gnirke A, Meissner A (April 2011). "Preparation of reduced representation bisulfite sequencing libraries for genome-scale DNA methylation profiling". Nature Protocols. 6 (4): 468–81. doi:10.1038/nprot.2010.190. PMID 21412275. S2CID 24912438.
- Fu Y, Luo GZ, Chen K, Deng X, Yu M, Han D, et al. (May 2015). "N6-methyldeoxyadenosine marks active transcription start sites in Chlamydomonas". Cell. 161 (4): 879–892. doi:10.1016/j.cell.2015.04.010. PMC 4427561. PMID 25936837.
- Beaulaurier J, Schadt EE, Fang G (March 2019). "Deciphering bacterial epigenomes using modern sequencing technologies". Nature Reviews. Genetics. 20 (3): 157–172. doi:10.1038/s41576-018-0081-3. PMC 6555402. PMID 30546107.
- Guo H, Zhu P, Yan L, Li R, Hu B, Lian Y, et al. (July 2014). "The DNA methylation landscape of human early embryos". Nature. 511 (7511): 606–10. Bibcode:2014Natur.511..606G. doi:10.1038/nature13544. PMID 25079557. S2CID 4450377.
- Gkountela S, Castro-Giner F, Szczerba BM, Vetter M, Landin J, Scherrer R, et al. (January 2019). "Circulating Tumor Cell Clustering Shapes DNA Methylation to Enable Metastasis Seeding". Cell. 176 (1–2): 98–112.e14. doi:10.1016/j.cell.2018.11.046. PMC 6363966. PMID 30633912.
- Stein RA (1 Jul 2019). "Single-Cell Sequencing Sifts through Multiple Omics". Retrieved 1 August 2019.
- "Shapiro E, Biezuner T, Linnarsson S (September 2013). "Single-cell sequencing-based technologies will revolutionize whole-organism science". Nature Reviews. Genetics. 14 (9): 618–30. doi:10.1038/nrg3542. PMID 23897237. S2CID 500845."
- Kolodziejczyk AA, Kim JK, Svensson V, Marioni JC, Teichmann SA (May 2015). "The technology and biology of single-cell RNA sequencing". Molecular Cell. 58 (4): 610–20. doi:10.1016/j.molcel.2015.04.005. PMID 26000846.
- Tammela, Tuomas; Sage, Julien (2020). "Investigating Tumor Heterogeneity in Mouse Models". Annual Review of Cancer Biology. 4: 99–119. doi:10.1146/annurev-cancerbio-030419-033413.
- Harris, Chris (2020). Single Cell Transcriptome Analysis in Prostate Cancer (MSc). University of Otago.
- Montoro DT, Haber AL, Biton M, Vinarsky V, Lin B, Birket SE, et al. (August 2018). "A revised airway epithelial hierarchy includes CFTR-expressing ionocytes". Nature. 560 (7718): 319–324. Bibcode:2018Natur.560..319M. doi:10.1038/s41586-018-0393-7. PMC 6295155. PMID 30069044.
- Plasschaert LW, Žilionis R, Choo-Wing R, Savova V, Knehr J, Roma G, et al. (August 2018). "A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte". Nature. 560 (7718): 377–381. Bibcode:2018Natur.560..377P. doi:10.1038/s41586-018-0394-6. PMC 6108322. PMID 30069046.
- Klein AM, Mazutis L, Akartuna I, Tallapragada N, Veres A, Li V, et al. (May 2015). "Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells". Cell. 161 (5): 1187–1201. doi:10.1016/j.cell.2015.04.044. PMC 4441768. PMID 26000487.
- Macosko EZ, Basu A, Satija R, Nemesh J, Shekhar K, Goldman M, et al. (May 2015). "Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets". Cell. 161 (5): 1202–1214. doi:10.1016/j.cell.2015.05.002. PMC 4481139. PMID 26000488.
- "Hebenstreit D (November 2012). "Methods, Challenges and Potentials of Single Cell RNA-seq". Biology. 1 (3): 658–67. doi:10.3390/biology1030658. PMC 4009822. PMID 24832513."
- Tang F, Barbacioru C, Wang Y, Nordman E, Lee C, Xu N, et al. (May 2009). "mRNA-Seq whole-transcriptome analysis of a single cell". Nature Methods. 6 (5): 377–82. doi:10.1038/NMETH.1315. PMID 19349980. S2CID 16570747.
- Islam S, Kjällquist U, Moliner A, Zajac P, Fan JB, Lönnerberg P, Linnarsson S (July 2011). "Characterization of the single-cell transcriptional landscape by highly multiplex RNA-seq". Genome Research. 21 (7): 1160–7. doi:10.1101/gr.110882.110. PMC 3129258. PMID 21543516.
- Ramsköld D, Luo S, Wang YC, Li R, Deng Q, Faridani OR, et al. (August 2012). "Full-length mRNA-Seq from single-cell levels of RNA and individual circulating tumor cells". Nature Biotechnology. 30 (8): 777–82. doi:10.1038/nbt.2282. PMC 3467340. PMID 22820318.
- Hashimshony T, Wagner F, Sher N, Yanai I (September 2012). "CEL-Seq: single-cell RNA-Seq by multiplexed linear amplification". Cell Reports. 2 (3): 666–73. doi:10.1016/j.celrep.2012.08.003. PMID 22939981.
- Singh M, Al-Eryani G, Carswell S, Ferguson JM, Blackburn J, Barton K, et al. (July 2019). "High-throughput targeted long-read single cell sequencing reveals the clonal and transcriptional landscape of lymphocytes". Nature Communications. 10 (1): 3120. Bibcode:2019NatCo..10.3120S. doi:10.1038/s41467-019-11049-4. PMC 6635368. PMID 31311926.
- Sasagawa Y, Nikaido I, Hayashi T, Danno H, Uno KD, Imai T, Ueda HR (April 2013). "Quartz-Seq: a highly reproducible and sensitive single-cell RNA sequencing method, reveals non-genetic gene-expression heterogeneity". Genome Biology. 14 (4): R31. doi:10.1186/gb-2013-14-4-r31. PMC 4054835. PMID 23594475.
- Kouno T, Moody J, Kwon AT, Shibayama Y, Kato S, Huang Y, et al. (January 2019). "C1 CAGE detects transcription start sites and enhancer activity at single-cell resolution". Nature Communications. 10 (1): 360. Bibcode:2019NatCo..10..360K. doi:10.1038/s41467-018-08126-5. PMC 6341120. PMID 30664627.
- Dal Molin A, Di Camillo B (July 2019). "How to design a single-cell RNA-sequencing experiment: pitfalls, challenges and perspectives". Briefings in Bioinformatics. 20 (4): 1384–1394. doi:10.1093/bib/bby007. PMID 29394315.
- Peterson VM, Zhang KX, Kumar N, Wong J, Li L, Wilson DC, et al. (October 2017). "Multiplexed quantification of proteins and transcripts in single cells". Nature Biotechnology. 35 (10): 936–939. doi:10.1038/nbt.3973. PMID 28854175. S2CID 205285357.
- Stoeckius M, Hafemeister C, Stephenson W, Houck-Loomis B, Chattopadhyay PK, Swerdlow H, et al. (September 2017). "Simultaneous epitope and transcriptome measurement in single cells". Nature Methods. 14 (9): 865–868. doi:10.1038/nmeth.4380. PMC 5669064. PMID 28759029.
- Hagemann-Jensen M, Abdullayev I, Sandberg R, Faridani OR (October 2018). "Small-seq for single-cell small-RNA sequencing". Nature Protocols. 13 (10): 2407–2424. doi:10.1038/s41596-018-0049-y. PMID 30250291. S2CID 52813142.
- Hayashi T, Ozaki H, Sasagawa Y, Umeda M, Danno H, Nikaido I (February 2018). "Single-cell full-length total RNA sequencing uncovers dynamics of recursive splicing and enhancer RNAs". Nature Communications. 9 (1): 619. Bibcode:2018NatCo...9..619H. doi:10.1038/s41467-018-02866-0. PMC 5809388. PMID 29434199.
- Armour CD, Castle JC, Chen R, Babak T, Loerch P, Jackson S, et al. (September 2009). "Digital transcriptome profiling using selective hexamer priming for cDNA synthesis". Nature Methods. 6 (9): 647–9. doi:10.1038/nmeth.1360. PMID 19668204. S2CID 12164981.
- Griffiths, JA; Scialdone, A; Marioni, JC (16 April 2018). "Using single-cell genomics to understand developmental processes and cell fate decisions". Molecular Systems Biology. 14 (4): e8046. doi:10.15252/msb.20178046. PMC 5900446. PMID 29661792.
- Raj B, Wagner DE, McKenna A, Pandey S, Klein AM, Shendure J, et al. (June 2018). "Simultaneous single-cell profiling of lineages and cell types in the vertebrate brain". Nature Biotechnology. 36 (5): 442–450. doi:10.1038/nbt.4103. PMC 5938111. PMID 29608178.
- Olmos D, Arkenau HT, Ang JE, Ledaki I, Attard G, Carden CP, et al. (January 2009). "Circulating tumour cell (CTC) counts as intermediate end points in castration-resistant prostate cancer (CRPC): a single-centre experience". Annals of Oncology. 20 (1): 27–33. doi:10.1093/annonc/mdn544. PMID 18695026.
- Levitin HM, Yuan J, Sims PA (April 2018). "Single-Cell Transcriptomic Analysis of Tumor Heterogeneity". Trends in Cancer. 4 (4): 264–268. doi:10.1016/j.trecan.2018.02.003. PMC 5993208. PMID 29606308.
- Jerby-Arnon L, Shah P, Cuoco MS, Rodman C, Su MJ, Melms JC, et al. (November 2018). "A Cancer Cell Program Promotes T Cell Exclusion and Resistance to Checkpoint Blockade". Cell. 175 (4): 984–997.e24. doi:10.1016/j.cell.2018.09.006. PMC 6410377. PMID 30388455.
- Neu, KE; Tang, Q; Wilson, PC; Khan, AA (February 2017). "Single-Cell Genomics: Approaches and Utility in Immunology". Trends in Immunology. 38 (2): 140–149. doi:10.1016/j.it.2016.12.001. PMC 5479322. PMID 28094102.
- Stephenson W, Donlin LT, Butler A, Rozo C, Bracken B, Rashidfarrokhi A, et al. (February 2018). "Single-cell RNA-seq of rheumatoid arthritis synovial tissue using low-cost microfluidic instrumentation". Nature Communications. 9 (1): 791. Bibcode:2018NatCo...9..791S. doi:10.1038/s41467-017-02659-x. PMC 5824814. PMID 29476078.
- Kuppe, Christoph; Ibrahim, Mahmoud M.; Kranz, Jennifer; Zhang, Xiaoting; Ziegler, Susanne; Perales-Patón, Javier; Jansen, Jitske; Reimer, Katharina C.; Smith, James R.; Dobie, Ross; Wilson-Kanamari, John R. (2020-11-11). "Decoding myofibroblast origins in human kidney fibrosis". Nature: 1–9. doi:10.1038/s41586-020-2941-1. ISSN 1476-4687. PMID 33176333.
- Avraham R, Haseley N, Brown D, Penaranda C, Jijon HB, Trombetta JJ, et al. (September 2015). "Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses". Cell. 162 (6): 1309–21. doi:10.1016/j.cell.2015.08.027. PMC 4578813. PMID 26343579.
- Bossel Ben-Moshe N, Hen-Avivi S, Levitin N, Yehezkel D, Oosting M, Joosten LA, et al. (July 2019). "Predicting bacterial infection outcomes using single cell RNA-sequencing analysis of human immune cells". Nature Communications. 10 (1): 3266. Bibcode:2019NatCo..10.3266B. doi:10.1038/s41467-019-11257-y. PMC 6646406. PMID 31332193.
- Mercatelli, Daniele; Ray, Forest; Giorgi, Federico M. (2019). "Pan-Cancer and Single-Cell Modeling of Genomic Alterations Through Gene Expression". Frontiers in Genetics. 10: 671. doi:10.3389/fgene.2019.00671. ISSN 1664-8021. PMC 6657420. PMID 31379928.
- Reid AJ, Talman AM, Bennett HM, Gomes AR, Sanders MJ, Illingworth CJ, et al. (March 2018). "Single-cell RNA-seq reveals hidden transcriptional variation in malaria parasites". eLife. 7: e33105. doi:10.7554/eLife.33105. PMC 5871331. PMID 29580379.
- Poran A, Nötzel C, Aly O, Mencia-Trinchant N, Harris CT, Guzman ML, et al. (November 2017). "Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites". Nature. 551 (7678): 95–99. Bibcode:2017Natur.551...95P. doi:10.1038/nature24280. PMC 6055935. PMID 29094698.
- Gasch AP, Yu FB, Hose J, Escalante LE, Place M, Bacher R, et al. (December 2017). Balaban N (ed.). "Single-cell RNA sequencing reveals intrinsic and extrinsic regulatory heterogeneity in yeast responding to stress". PLOS Biology. 15 (12): e2004050. doi:10.1371/journal.pbio.2004050. PMC 5746276. PMID 29240790.
- Kang Y, Norris MH, Zarzycki-Siek J, Nierman WC, Donachie SP, Hoang TT (June 2011). "Transcript amplification from single bacterium for transcriptome analysis". Genome Research. 21 (6): 925–35. doi:10.1101/gr.116103.110. PMC 3106325. PMID 21536723.
- Cao J, Packer JS, Ramani V, Cusanovich DA, Huynh C, Daza R, et al. (August 2017). "Comprehensive single-cell transcriptional profiling of a multicellular organism". Science. 357 (6352): 661–667. Bibcode:2017Sci...357..661C. doi:10.1126/science.aam8940. PMC 5894354. PMID 28818938.
- Plass M, Solana J, Wolf FA, Ayoub S, Misios A, Glažar P, et al. (May 2018). "Cell type atlas and lineage tree of a whole complex animal by single-cell transcriptomics". Science. 360 (6391): eaaq1723. doi:10.1126/science.aaq1723. PMID 29674432.
- Fincher CT, Wurtzel O, de Hoog T, Kravarik KM, Reddien PW (May 2018). "Schmidtea mediterranea". Science. 360 (6391): eaaq1736. doi:10.1126/science.aaq1736. PMC 6563842. PMID 29674431.
- Gerber, Tobias; Murawala, Prayag; Knapp, Dunja; Masselink, Wouter; Schuez, Maritta; Hermann, Sarah; Gac-Santel, Malgorzata; Nowoshilow, Sergej; Kageyama, Jorge; Khattak, Shahryar; Currie, Joshua D. (2018-10-26). "Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration". Science. 362 (6413): eaaq0681. Bibcode:2018Sci...362..681G. doi:10.1126/science.aaq0681. ISSN 0036-8075. PMC 6669047. PMID 30262634.
- Leigh, Nicholas D.; Dunlap, Garrett S.; Johnson, Kimberly; Mariano, Rachelle; Oshiro, Rachel; Wong, Alan Y.; Bryant, Donald M.; Miller, Bess M.; Ratner, Alex; Chen, Andy; Ye, William W. (2018-12-04). "Transcriptomic landscape of the blastema niche in regenerating adult axolotl limbs at single-cell resolution". Nature Communications. 9 (1): 5153. Bibcode:2018NatCo...9.5153L. doi:10.1038/s41467-018-07604-0. ISSN 2041-1723. PMC 6279788. PMID 30514844.
- Wagner DE, Weinreb C, Collins ZM, Briggs JA, Megason SG, Klein AM (June 2018). "Single-cell mapping of gene expression landscapes and lineage in the zebrafish embryo". Science. 360 (6392): 981–987. Bibcode:2018Sci...360..981W. doi:10.1126/science.aar4362. PMC 6083445. PMID 29700229.
- Farrell JA, Wang Y, Riesenfeld SJ, Shekhar K, Regev A, Schier AF (June 2018). "Single-cell reconstruction of developmental trajectories during zebrafish embryogenesis". Science. 360 (6392): eaar3131. doi:10.1126/science.aar3131. PMC 6247916. PMID 29700225.
- Zafar H, Lin C, Bar-Joseph Z (June 2020). "Single-cell lineage tracing by integrating CRISPR-Cas9 mutations with transcriptomic data". Nature Communications. 3055 (1): 3055. Bibcode:2020NatCo..11.3055Z. doi:10.1038/s41467-020-16821-5. PMC 7298005. PMID 32546686.
- Briggs JA, Weinreb C, Wagner DE, Megason S, Peshkin L, Kirschner MW, Klein AM (June 2018). "The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution". Science. 360 (6392): eaar5780. doi:10.1126/science.aar5780. PMC 6038144. PMID 29700227.
- You J. "Science's 2018 Breakthrough of the Year: tracking development cell by cell". Science Magazine. American Association for the Advancement of Science.
- Kurimoto K, Yabuta Y, Ohinata Y, Saitou M (2007). "Global single-cell cDNA amplification to provide a template for representative high-density oligonucleotide microarray analysis". Nature Protocols. 2 (3): 739–52. doi:10.1038/nprot.2007.79. PMID 17406636. S2CID 7404545.
- Yachida S, Jones S, Bozic I, Antal T, Leary R, Fu B, et al. (October 2010). "Distant metastasis occurs late during the genetic evolution of pancreatic cancer". Nature. 467 (7319): 1114–7. Bibcode:2010Natur.467.1114Y. doi:10.1038/nature09515. PMC 3148940. PMID 20981102.
- Frumkin D, Wasserstrom A, Itzkovitz S, Harmelin A, Rechavi G, Shapiro E (February 2008). "Amplification of multiple genomic loci from single cells isolated by laser micro-dissection of tissues". BMC Biotechnology. 8 (17): 17. doi:10.1186/1472-6750-8-17. PMC 2266725. PMID 18284708.
- Dalerba P, Kalisky T, Sahoo D, Rajendran PS, Rothenberg ME, Leyrat AA, et al. (November 2011). "Single-cell dissection of transcriptional heterogeneity in human colon tumors". Nature Biotechnology. 29 (12): 1120–7. doi:10.1038/nbt.2038. PMC 3237928. PMID 22081019.
- White AK, VanInsberghe M, Petriv OI, Hamidi M, Sikorski D, Marra MA, et al. (August 2011). "High-throughput microfluidic single-cell RT-qPCR". Proceedings of the National Academy of Sciences of the United States of America. 108 (34): 13999–4004. Bibcode:2011PNAS..10813999W. doi:10.1073/pnas.1019446108. PMC 3161570. PMID 21808033.