YAP1
YAP1 (yes-associated protein 1), also known as YAP or YAP65, is a protein that acts as a transcriptional regulator by activating the transcription of genes involved in cell proliferation and suppressing apoptotic genes. YAP1 is inhibited in the Hippo signaling pathway which allows the cellular control of organ size and tumor suppression. YAP1 was first identified by virtue of its ability to associate with the SH3 domain of Yes and Src protein tyrosine kinases.[5] YAP1 is a potent oncogene, which is amplified in various human cancers.[6][7]




Structure
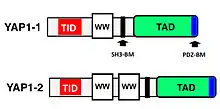
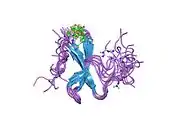
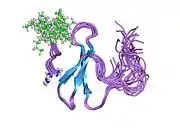
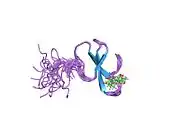
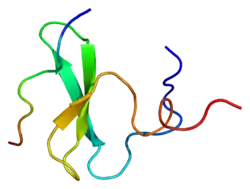
Cloning of the YAP1 gene facilitated the identification of a modular protein domain, known as the WW domain.[8][9][10] Two splice isoforms of the YAP1 gene product were initially identified, named YAP1-1 and YAP1-2, which differed by the presence of an extra 38 amino acids that encoded the WW domain.[11][12] Apart from the WW domain, the modular structure of YAP1 contains a proline-rich region at the very amino terminus, which is followed by a TID (TEAD transcription factor interacting domain).[13] Next, following a single WW domain, which is present in the YAP1-1 isoform, and two WW domains, which are present in the YAP1-2 isoform, there is the SH3-BM (Src Homology 3 binding motif).[5][14] Following the SH3-BM is a TAD (transcription activation domain) and a PDZ domain-binding motif (PDZ-BM) (Figure 1).[15][16]
Function
YAP1 is a transcriptional co-activator[17] and its proliferative and oncogenic activity is driven by its association with the TEAD family of transcription factors,[13] which up-regulate genes that promote cell growth and inhibit apoptosis.[18] Several other functional partners of YAP1 were identified, including RUNX,[17] SMADs,[19][20] p73,[21] ErbB4,[22][23] TP53BP,[24] LATS1/2,[25] PTPN14,[26] AMOTs,[27][28][29][30] and ZO1/2.[31] YAP1 and its close paralog, TAZ (WWTR1), are the main effectors of the Hippo tumor suppressor pathway.[32] When the pathway is activated, YAP1 and TAZ are phosphorylated on a serine residue and sequestered in the cytoplasm by 14-3-3 proteins.[32] When the Hippo pathway is not activated, YAP1/TAZ enter the nucleus and regulate gene expression.[32]
It is reported that several genes are regulated by YAP1, including Birc2, Birc5, connective tissue growth factor (CTGF), amphiregulin (AREG), Cyr61, Hoxa1 and Hoxc13.
YAP/TAZ have also been shown to act as stiffness sensors, regulating mechanotransduction independently of the Hippo signalling cascade.[33]
As YAP and TAZ are transcriptional co-activators, they do not have DNA-binding domains. Instead, when inside the nucleus, they regulate gene expression through TEAD1-4 which are sequence-specific transcription factors that mediate the main transcriptional output of the Hippo pathway.[34] The YAP/TAZ and TEAD interaction competitively inhibits and actively dissociates the TEAD/VGLL4 interaction which functions as a transcriptional repressor.[35] Mouse models with YAP over-expression have been shown to exhibit up-regulation of the TEAD target gene expression which results in increased expansion of progenitor cells and tissue overgrowth.[36]
Regulation
Biochemical Regulation
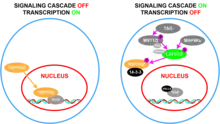
At the biochemical level, YAP is part of and regulated by the Hippo signaling pathway where a kinase cascade results in its “inactivation”, along with that of TAZ.[37] In this signaling cascade, TAO kinases phosphorylate Ste20-like kinases, MST1/2, at their activation loops (Thr183 for MST1 and Thr180 for MST2).[38][39] Active MST1/2 then phosphorylate SAV1 and MOB1A/B which are scaffold proteins that assist in the recruitment and phosphorylation of LATS1/2.[40][41] LATS1/2 can also be phosphorylated by two groups of MAP4Ks.[42][43] LATS1/2 then phosphorylate YAP and TAZ which causes them to bind with 14-3-3, resulting in cytoplasmic sequestration of YAP and TAZ.[44] The result of the activation of this pathway is the restriction of YAP/TAZ from entering the cell nucleus.
Mechanotransductive Regulation
Additionally, YAP is regulated by mechanical cues such as extracellular matrix (ECM) rigidity, strain, shear stress, or adhesive area, processes that are reliant on cytoskeletal integrity.[45] These mechanically induced localization phenomena are thought to be the result of nuclear flattening induced pore size change, mechanosensitive nuclear membrane ion channels, mechanical protein stability, or a variety of other factors.[45] These mechanical factors have also been linked to certain cancer cells via nuclear softening and higher ECM stiffnesses.[46][47][48] Under this framework, the nuclear softening phenotype of cancer cells would promote nuclear flattening in response to a force, causing YAP localization, which could explain its over-expression and promoted proliferation in oncogenic cells.[49] Additionally, the higher ECM stiffness phenotype commonly seen in tumors due to enhanced integrin signaling,[48] could flatten the cell and nucleus, once again causing higher YAP nuclear localization. Likewise, the opposite effect of nuclear stiffening as a result of a variety of stimuli such as an over-expression of lamin A, has been shown to decrease nuclear YAP localization.[50][51]
Oncogenic Role
Recent findings on the role of the Hippo signaling pathway in cancer progression has spurred great anticipation and interest.[52] Hyperactivation of YAP and TAZ has been pervasively observed amongst many cancers and YAP/TAZ-mediated transcriptional activity is implicated in the development of abnormal cell growth.[49][53][54] However, while YAP has been identified as a proto-oncogene, recent studies have also shown its function as a tumor suppressor depending on cellular context.[55][56]
Clinical Significance
Heterozygous loss-of-function mutations in the YAP1 gene have been identified in two families with major eye malformations with or without extra-ocular features such as hearing loss, cleft lip, intellectual disability and renal disease.[57]
The YAP1 oncogene serves as a target for the development of new cancer drugs.[58] Small compounds have been identified that disrupt the YAP1-TEAD complex or block the binding function of WW domains.[59][60] These small molecules represent lead compounds for the development of therapies for cancer patients, who harbor amplified or overexpressed YAP oncogene.
The Hippo/YAP signaling pathway may exert neuroprotective effects through mitigating blood-brain barrier disruption after cerebral ischemia/reperfusion injury.[61]
External links
References
- GRCh38: Ensembl release 89: ENSG00000137693 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000053110 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Sudol M (August 1994). "Yes-associated protein (YAP65) is a proline-rich phosphoprotein that binds to the SH3 domain of the Yes proto-oncogene product". Oncogene. 9 (8): 2145–52. PMID 8035999.
- Huang J, Wu S, Barrera J, Matthews K, Pan D (August 2005). "The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP". Cell. 122 (3): 421–34. doi:10.1016/j.cell.2005.06.007. PMID 16096061. S2CID 14139806.
- Overholtzer M, Zhang J, Smolen GA, Muir B, Li W, Sgroi DC, et al. (August 2006). "Transforming properties of YAP, a candidate oncogene on the chromosome 11q22 amplicon". Proceedings of the National Academy of Sciences of the United States of America. 103 (33): 12405–10. Bibcode:2006PNAS..10312405O. doi:10.1073/pnas.0605579103. PMC 1533802. PMID 16894141.
- Bork P, Sudol M (December 1994). "The WW domain: a signalling site in dystrophin?". Trends in Biochemical Sciences. 19 (12): 531–3. doi:10.1016/0968-0004(94)90053-1. PMID 7846762.
- André B, Springael JY (December 1994). "WWP, a new amino acid motif present in single or multiple copies in various proteins including dystrophin and the SH3-binding Yes-associated protein YAP65". Biochemical and Biophysical Research Communications. 205 (2): 1201–5. doi:10.1006/bbrc.1994.2793. PMID 7802651.
- Hofmann K, Bucher P (January 1995). "The rsp5-domain is shared by proteins of diverse functions". FEBS Letters. 358 (2): 153–7. doi:10.1016/0014-5793(94)01415-W. PMID 7828727. S2CID 23110605.
- Sudol M, Bork P, Einbond A, Kastury K, Druck T, Negrini M, et al. (June 1995). "Characterization of the mammalian YAP (Yes-associated protein) gene and its role in defining a novel protein module, the WW domain". The Journal of Biological Chemistry. 270 (24): 14733–41. doi:10.1074/jbc.270.24.14733. PMID 7782338.
- Gaffney CJ, Oka T, Mazack V, Hilman D, Gat U, Muramatsu T, et al. (November 2012). "Identification, basic characterization and evolutionary analysis of differentially spliced mRNA isoforms of human YAP1 gene". Gene. 509 (2): 215–22. doi:10.1016/j.gene.2012.08.025. PMC 3455135. PMID 22939869.
- Vassilev A, Kaneko KJ, Shu H, Zhao Y, DePamphilis ML (May 2001). "TEAD/TEF transcription factors utilize the activation domain of YAP65, a Src/Yes-associated protein localized in the cytoplasm". Genes & Development. 15 (10): 1229–41. doi:10.1101/gad.888601. PMC 313800. PMID 11358867.
- Ren R, Mayer BJ, Cicchetti P, Baltimore D (February 1993). "Identification of a ten-amino acid proline-rich SH3 binding site". Science. 259 (5098): 1157–61. Bibcode:1993Sci...259.1157R. doi:10.1126/science.8438166. PMID 8438166.
- Wang S, Raab RW, Schatz PJ, Guggino WB, Li M (May 1998). "Peptide binding consensus of the NHE-RF-PDZ1 domain matches the C-terminal sequence of cystic fibrosis transmembrane conductance regulator (CFTR)". FEBS Letters. 427 (1): 103–8. doi:10.1016/S0014-5793(98)00402-5. PMID 9613608. S2CID 20803242.
- Mohler PJ, Kreda SM, Boucher RC, Sudol M, Stutts MJ, Milgram SL (November 1999). "Yes-associated protein 65 localizes p62(c-Yes) to the apical compartment of airway epithelia by association with EBP50". The Journal of Cell Biology. 147 (4): 879–90. doi:10.1083/jcb.147.4.879. PMC 2156157. PMID 10562288.
- Yagi R, Chen LF, Shigesada K, Murakami Y, Ito Y (May 1999). "A WW domain-containing yes-associated protein (YAP) is a novel transcriptional co-activator". The EMBO Journal. 18 (9): 2551–62. doi:10.1093/emboj/18.9.2551. PMC 1171336. PMID 10228168.
- Zhao B, Kim J, Ye X, Lai ZC, Guan KL (February 2009). "Both TEAD-binding and WW domains are required for the growth stimulation and oncogenic transformation activity of yes-associated protein". Cancer Research. 69 (3): 1089–98. doi:10.1158/0008-5472.CAN-08-2997. PMID 19141641.
- Ferrigno O, Lallemand F, Verrecchia F, L'Hoste S, Camonis J, Atfi A, Mauviel A (July 2002). "Yes-associated protein (YAP65) interacts with Smad7 and potentiates its inhibitory activity against TGF-beta/Smad signaling". Oncogene. 21 (32): 4879–84. doi:10.1038/sj.onc.1205623. PMID 12118366.
- Aragón E, Goerner N, Xi Q, Gomes T, Gao S, Massagué J, Macias MJ (October 2012). "Structural basis for the versatile interactions of Smad7 with regulator WW domains in TGF-β Pathways". Structure. 20 (10): 1726–36. doi:10.1016/j.str.2012.07.014. PMC 3472128. PMID 22921829.
- Strano S, Munarriz E, Rossi M, Castagnoli L, Shaul Y, Sacchi A, et al. (May 2001). "Physical interaction with Yes-associated protein enhances p73 transcriptional activity". The Journal of Biological Chemistry. 276 (18): 15164–73. doi:10.1074/jbc.M010484200. PMID 11278685.
- Komuro A, Nagai M, Navin NE, Sudol M (August 2003). "WW domain-containing protein YAP associates with ErbB-4 and acts as a co-transcriptional activator for the carboxyl-terminal fragment of ErbB-4 that translocates to the nucleus". The Journal of Biological Chemistry. 278 (35): 33334–41. doi:10.1074/jbc.M305597200. PMID 12807903.
- Omerovic J, Puggioni EM, Napoletano S, Visco V, Fraioli R, Frati L, et al. (April 2004). "Ligand-regulated association of ErbB-4 to the transcriptional co-activator YAP65 controls transcription at the nuclear level". Experimental Cell Research. 294 (2): 469–79. doi:10.1016/j.yexcr.2003.12.002. PMID 15023535.
- Espanel X, Sudol M (April 2001). "Yes-associated protein and p53-binding protein-2 interact through their WW and SH3 domains". The Journal of Biological Chemistry. 276 (17): 14514–23. doi:10.1074/jbc.M008568200. PMID 11278422.
- Oka T, Mazack V, Sudol M (October 2008). "Mst2 and Lats kinases regulate apoptotic function of Yes kinase-associated protein (YAP)". The Journal of Biological Chemistry. 283 (41): 27534–46. doi:10.1074/jbc.M804380200. PMID 18640976.
- Liu X, Yang N, Figel SA, Wilson KE, Morrison CD, Gelman IH, Zhang J (March 2013). "PTPN14 interacts with and negatively regulates the oncogenic function of YAP". Oncogene. 32 (10): 1266–73. doi:10.1038/onc.2012.147. PMC 4402938. PMID 22525271.
- Wang W, Huang J, Chen J (February 2011). "Angiomotin-like proteins associate with and negatively regulate YAP1". The Journal of Biological Chemistry. 286 (6): 4364–70. doi:10.1074/jbc.C110.205401. PMC 3039387. PMID 21187284.
- Chan SW, Lim CJ, Chong YF, Pobbati AV, Huang C, Hong W (March 2011). "Hippo pathway-independent restriction of TAZ and YAP by angiomotin". The Journal of Biological Chemistry. 286 (9): 7018–26. doi:10.1074/jbc.C110.212621. PMC 3044958. PMID 21224387.
- Zhao B, Li L, Lu Q, Wang LH, Liu CY, Lei Q, Guan KL (January 2011). "Angiomotin is a novel Hippo pathway component that inhibits YAP oncoprotein". Genes & Development. 25 (1): 51–63. doi:10.1101/gad.2000111. PMC 3012936. PMID 21205866.
- Oka T, Schmitt AP, Sudol M (January 2012). "Opposing roles of angiomotin-like-1 and zona occludens-2 on pro-apoptotic function of YAP". Oncogene. 31 (1): 128–34. doi:10.1038/onc.2011.216. PMID 21685940.
- Oka T, Remue E, Meerschaert K, Vanloo B, Boucherie C, Gfeller D, et al. (December 2010). "Functional complexes between YAP2 and ZO-2 are PDZ domain-dependent, and regulate YAP2 nuclear localization and signalling". The Biochemical Journal (Submitted manuscript). 432 (3): 461–72. doi:10.1042/BJ20100870. hdl:1854/LU-1256657. PMID 20868367.
- Pan D (October 2010). "The hippo signaling pathway in development and cancer". Developmental Cell. 19 (4): 491–505. doi:10.1016/j.devcel.2010.09.011. PMC 3124840. PMID 20951342.
- McMurray RJ, Dalby MJ, Tsimbouri PM (May 2015). "Using biomaterials to study stem cell mechanotransduction, growth and differentiation" (PDF). Journal of Tissue Engineering and Regenerative Medicine. 9 (5): 528–39. doi:10.1002/term.1957. PMID 25370612. S2CID 39642567.
- Zhao B, Ye X, Yu J, Li L, Li W, Li S, et al. (July 2008). "TEAD mediates YAP-dependent gene induction and growth control". Genes & Development. 22 (14): 1962–71. doi:10.1101/gad.1664408. PMC 2492741. PMID 18579750.
- Koontz LM, Liu-Chittenden Y, Yin F, Zheng Y, Yu J, Huang B, et al. (May 2013). "The Hippo effector Yorkie controls normal tissue growth by antagonizing scalloped-mediated default repression". Developmental Cell. 25 (4): 388–401. doi:10.1016/j.devcel.2013.04.021. PMC 3705890. PMID 23725764.
- Chen Q, Zhang N, Xie R, Wang W, Cai J, Choi KS, et al. (June 2015). "Homeostatic control of Hippo signaling activity revealed by an endogenous activating mutation in YAP". Genes & Development. 29 (12): 1285–97. doi:10.1101/gad.264234.115. PMC 4495399. PMID 26109051.
- Meng Z, Moroishi T, Guan KL (January 2016). "Mechanisms of Hippo pathway regulation". Genes & Development. 30 (1): 1–17. doi:10.1101/gad.274027.115. PMC 4701972. PMID 26728553.
- Boggiano JC, Vanderzalm PJ, Fehon RG (November 2011). "Tao-1 phosphorylates Hippo/MST kinases to regulate the Hippo-Salvador-Warts tumor suppressor pathway". Developmental Cell. 21 (5): 888–95. doi:10.1016/j.devcel.2011.08.028. PMC 3217187. PMID 22075147.
- Poon CL, Lin JI, Zhang X, Harvey KF (November 2011). "The sterile 20-like kinase Tao-1 controls tissue growth by regulating the Salvador-Warts-Hippo pathway". Developmental Cell. 21 (5): 896–906. doi:10.1016/j.devcel.2011.09.012. PMID 22075148.
- Callus BA, Verhagen AM, Vaux DL (September 2006). "Association of mammalian sterile twenty kinases, Mst1 and Mst2, with hSalvador via C-terminal coiled-coil domains, leads to its stabilization and phosphorylation". The FEBS Journal. 273 (18): 4264–76. doi:10.1111/j.1742-4658.2006.05427.x. PMID 16930133. S2CID 8261982.
- Praskova M, Xia F, Avruch J (March 2008). "MOBKL1A/MOBKL1B phosphorylation by MST1 and MST2 inhibits cell proliferation". Current Biology. 18 (5): 311–21. doi:10.1016/j.cub.2008.02.006. PMC 4682548. PMID 18328708.
- Meng Z, Moroishi T, Mottier-Pavie V, Plouffe SW, Hansen CG, Hong AW, et al. (October 2015). "MAP4K family kinases act in parallel to MST1/2 to activate LATS1/2 in the Hippo pathway". Nature Communications. 6: 8357. Bibcode:2015NatCo...6.8357M. doi:10.1038/ncomms9357. PMC 4600732. PMID 26437443.
- Zheng Y, Wang W, Liu B, Deng H, Uster E, Pan D (September 2015). "Identification of Happyhour/MAP4K as Alternative Hpo/Mst-like Kinases in the Hippo Kinase Cascade". Developmental Cell. 34 (6): 642–55. doi:10.1016/j.devcel.2015.08.014. PMC 4589524. PMID 26364751.
- Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim J, et al. (November 2007). "Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control". Genes & Development. 21 (21): 2747–61. doi:10.1101/gad.1602907. PMC 2045129. PMID 17974916.
- Elosegui-Artola A, Andreu I, Beedle AE, Lezamiz A, Uroz M, Kosmalska AJ, et al. (November 2017). "Force Triggers YAP Nuclear Entry by Regulating Transport across Nuclear Pores". Cell. 171 (6): 1397–1410.e14. doi:10.1016/j.cell.2017.10.008. PMID 29107331.
- Cross SE, Jin YS, Rao J, Gimzewski JK (December 2007). "Nanomechanical analysis of cells from cancer patients". Nature Nanotechnology. 2 (12): 780–3. Bibcode:2007NatNa...2..780C. doi:10.1038/nnano.2007.388. PMID 18654431.
- Guck J, Schinkinger S, Lincoln B, Wottawah F, Ebert S, Romeyke M, et al. (May 2005). "Optical deformability as an inherent cell marker for testing malignant transformation and metastatic competence". Biophysical Journal. 88 (5): 3689–98. Bibcode:2005BpJ....88.3689G. doi:10.1529/biophysj.104.045476. PMC 1305515. PMID 15722433.
- Friedl P, Alexander S (November 2011). "Cancer invasion and the microenvironment: plasticity and reciprocity". Cell. 147 (5): 992–1009. doi:10.1016/j.cell.2011.11.016. PMID 22118458.
- Shimomura T, Miyamura N, Hata S, Miura R, Hirayama J, Nishina H (January 2014). "The PDZ-binding motif of Yes-associated protein is required for its co-activation of TEAD-mediated CTGF transcription and oncogenic cell transforming activity". Biochemical and Biophysical Research Communications. 443 (3): 917–23. doi:10.1016/j.bbrc.2013.12.100. PMID 24380865.
- Swift J, Ivanovska IL, Buxboim A, Harada T, Dingal PC, Pinter J, et al. (August 2013). "Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation". Science. 341 (6149): 1240104. doi:10.1126/science.1240104. PMC 3976548. PMID 23990565.
- Gjorevski N, Sachs N, Manfrin A, Giger S, Bragina ME, Ordóñez-Morán P, et al. (November 2016). "Designer matrices for intestinal stem cell and organoid culture". Nature. 539 (7630): 560–564. doi:10.1038/nature20168. PMID 27851739. S2CID 4470849.
- Moroishi T, Hansen CG, Guan KL (February 2015). "The emerging roles of YAP and TAZ in cancer". Nature Reviews. Cancer. 15 (2): 73–79. doi:10.1038/nrc3876. PMC 4562315. PMID 25592648.
- Harvey KF, Zhang X, Thomas DM (April 2013). "The Hippo pathway and human cancer". Nature Reviews. Cancer. 13 (4): 246–57. doi:10.1038/nrc3458. PMID 23467301. S2CID 2008641.
- Johnson R, Halder G (January 2014). "The two faces of Hippo: targeting the Hippo pathway for regenerative medicine and cancer treatment". Nature Reviews. Drug Discovery. 13 (1): 63–79. doi:10.1038/nrd4161. PMC 4167640. PMID 24336504.
- Barry ER, Morikawa T, Butler BL, Shrestha K, de la Rosa R, Yan KS, et al. (January 2013). "Restriction of intestinal stem cell expansion and the regenerative response by YAP". Nature. 493 (7430): 106–10. Bibcode:2013Natur.493..106B. doi:10.1038/nature11693. PMC 3536889. PMID 23178811.
- Cottini F, Hideshima T, Xu C, Sattler M, Dori M, Agnelli L, et al. (June 2014). "Rescue of Hippo coactivator YAP1 triggers DNA damage-induced apoptosis in hematological cancers". Nature Medicine. 20 (6): 599–606. doi:10.1038/nm.3562. PMC 4057660. PMID 24813251.
- Williamson KA, Rainger J, Floyd JA, Ansari M, Meynert A, Aldridge KV, et al. (February 2014). "Heterozygous loss-of-function mutations in YAP1 cause both isolated and syndromic optic fissure closure defects". American Journal of Human Genetics. 94 (2): 295–302. doi:10.1016/j.ajhg.2014.01.001. PMC 3928658. PMID 24462371.
- Sudol M, Shields DC, Farooq A (September 2012). "Structures of YAP protein domains reveal promising targets for development of new cancer drugs". Seminars in Cell & Developmental Biology. 23 (7): 827–33. doi:10.1016/j.semcdb.2012.05.002. PMC 3427467. PMID 22609812.
- Liu-Chittenden Y, Huang B, Shim JS, Chen Q, Lee SJ, Anders RA, et al. (June 2012). "Genetic and pharmacological disruption of the TEAD-YAP complex suppresses the oncogenic activity of YAP". Genes & Development. 26 (12): 1300–5. doi:10.1101/gad.192856.112. PMC 3387657. PMID 22677547.
- Kang SG, Huynh T, Zhou R (2012). "Non-destructive inhibition of metallofullerenol Gd@C(82)(OH)(22) on WW domain: implication on signal transduction pathway". Scientific Reports. 2: 957. Bibcode:2012NatSR...2E.957K. doi:10.1038/srep00957. PMC 3518810. PMID 23233876.
- Gong P, Zhang Z, Zou C, Tian Q, Chen X, Hong M, et al. (January 2019). "Hippo/YAP signaling pathway mitigates blood-brain barrier disruption after cerebral ischemia/reperfusion injury". Behavioural Brain Research. 356: 8–17. doi:10.1016/j.bbr.2018.08.003. PMC 6193462. PMID 30092249.
PDB gallery | |
|---|---|
|