Paleobiota of the Ciechocinek Formation
The Ciechocinek Formation is a Jurassic (lower to middle Toarcian) geologic formation which extends across the Baltic coast from Grimmen, Germany, to Nida, Lithuania, with its major sequence in Poland and boreholes in Kaliningrad. Dinosaur species uncovered here include the Thyreorporan Emausaurus and others with an unclassified genus.

| Part of a series on |
| Paleontology |
|---|
 |
|
Paleontology Portal Category |
In Poland, the main basin lacks marine microfauna. The Ciechocinek Formation in the Częstochowa-Zawiercie area reveals the remains of a wide range of prehistoric environments; the Fore-Sudetic Monocline region must have been an extensive bay similar to Lake Maracaibo in Venezuela. The basin's shore zone was a flat, muddy, marshy coastal plain.
The region has the remains of the Wrêczyca River, which was active for most of the Pliensbachian/Toarcian period. At the Brody-Lubienia borehole (Lubienia), which once formed part of the river's east side, an alluvial system ended at a delta and discharged into a shallow marine bay and lagoon. A number of phyllopods and fossilized plant roots have been found here, where they were discharged by the river. Paleosol indicates that the lagoon had a maximum depth of about 6 metres (20 ft).
Foraminifera
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ammobaculites[1] |
|
|
Cunchs |
A marine Foraminiferan, member of the family Ammomarginulininae inside Lituolidae. |
|
|
Placopsilina[1] |
|
|
Cunchs |
A marine Foraminiferan, member of the family Lituoloidea inside Lituolida. |
|
|
|
Cunchs |
A marine Foraminiferan, member of the family Lituoloidea inside Lituolida. |
||
|
lnvolutina[1] |
|
|
Cunchs |
A marine Foraminiferan, member of the family Ammodiscinae inside Ammodiscina. |
|
|
|
Cunchs |
A marine Foraminiferan, type member of the family Ammodiscinae inside Ammodiscina. Benthic foraminiferal successions heavily dominated by Ammodiscus and Trochammina are known from several Late Triassic to Middle Jurassic sediment packages along the Atlantic margin of northwestern Europe.[6] |
||
|
|
Cunchs |
A marine Foraminiferan, member of the family Usbekistaniinae inside Ammodiscidae. |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Trochammininae inside Trochamminina. Small-sized Ammodiscus–Trochammina assemblages are found associated with delta-influenced shelf environment, where biota would have been stressed by intermittent periods with moderate hypoxia combined with lowered salinity and storm impacts.[6] |
||
|
|
Cunchs |
A marine Foraminiferan, member of the family Saccamminidae inside Astrorhizacea. Sometimes confused with the genus Arlagenammum.[6] |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Saccamminidae inside Astrorhizacea. Sometimes confused with the genus Arsaccammum or called Saccamina.[6] The Local specimens are rather rare, incomplete and complicate to identify. |
||
|
Crithionina[1] |
|
|
Cunchs |
A marine Foraminiferan, type member of the family Saccamminidae inside Astrorhizacea. |
|
|
|
Cunchs |
A marine Foraminiferan, member of the family Vaginulinidae inside Nodosarioidea. |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Vaginulininae inside Nodosarioidea. |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Ophthalmidiidae inside Cornuspiroidea. |
||
|
|
Cunchs |
A marine Foraminiferan, member of the family Nodosariidae inside Nodosariinae. |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Lingulininae inside Nodosariinae. |
||
|
|
Cunchs |
A marine Foraminiferan, type member of the family Bolivinidae inside Bolivinoidea. |
||
Dinoflagellates
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Nannoceratopsis[10] |
|
|
Dinocysts |
A marine Dinophyceae Dinoflagellatan, type member of the family Nannoceratopsiaceae inside Nannoceratopsiales. The large amount of Cysts of the genus point to more diversified marine palaeoenvironments. N. senex is the most abundant. Locally Nannoceratopsis recovers series of marine transgressions and regressions, pointing to the presence of interbedding marine and brackish sediments. |
|
|
Luehndea[10] |
|
|
Dinocysts |
A marine Dinophyceae Dinoflagellatan, type member of the family Luehndeoideae inside Mancodiniaceae. Presence of Luehndea spinosa suggests Late Pliensbachian–earliest Toarcian age of studied assemblages. The marine dinoflagellate cyst Luehndea spinosa and foraminiferal linings were found only in the lower half of the Kozłowice succession. |
|
Fungi
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Saprophyte fungal Spores from Marine and Deltaic Settings associated with wood and Litter.[11] The frequency of fungal spores on the Polish basin on the Pliensbachian-Toarcian transition is correlated with negative C isotope peaks and enhanced cuticular plant litter accumulation, pointing to the climate-driven enhanced decomposition of wood and rapid destruction of terrestrial carbon, which may have played an important role in the aggravation of the Jurassic Greenhouse disaster.[11] Associated with a high rate of organic burial, the presence of Fungal Matter increased on the Uppermost layers of the Drzewica Formation, with a continued deposition between the T-OAE extinction, and several ups and downs on the Ciechocinek Formation, related with local climate and humidity changes. This is rather a reflection of the efficiency of terrestrial biodegradation.[11] Measured increasing of temperature favored local fungal-mediated decomposition of plant litter, specifically of normally resistant wood.[11] Observed fungal spores represent various morphotypes and resemble superficially other palynogenic detritus, such as spherical Prasinophyceae.[11] In some cases spherical fungal spores show structures related to Hyphae attachments.[11] |
.jpg.webp) Spores found on the Polish are mostly from Saprophyte (wood decomposers) fungi, resembling the extant genera like Scutellinia, which are probably the main origin for the Fungal Spores found on the Ciechocinek Formation | |
|
Amerospores[11] |
|
|
Fungal Spores |
A Saprophyte fungus, member of the family Sordariomycetes inside Ascomycota.[11] Non-filamentous spores with no septations and with no projections longer than the spore body. Related with the extant genus Poronia. Fungal spore peaks linked to a relative and absolute loss of wood suggest a prominent role of fungal wood decomposers.[11] |
_Fr_587104.jpg.webp) Poronia punctata, an extant saprophyte fungus with Amerospores. Similar fungi probably liberated this spores on the Ciechocinek Formation |
|
Phragmospores[11] |
|
|
Fungal Spores |
A Saprophyte fungus, member of the family Dothideomycetes or Sordariomycetes inside Ascomycota.[11] Spores with two or more transverse septa. Related with the extant genus Acanthostigma, facultative pathogen, or beneficial partner of many plant species. Local humidity, though sufficiently high for fungal development throughout the whole T-OAE interval, is therefore thought to have been of a lesser significance for dynamics of fungal decomposition than elevated temperature.[11] |
 Leptosphaeria acuta, an extant wood fungus with phragmospores |
|
Dictyosporiaceae[11] |
|
|
Fungal Spores |
A freshwater anamorphic fungus, member of the family Pleosporales inside Pleosporomycetidae.[11] Multicellular spore with septations that intersect in more than one plane. Characterized by being mostly aquatic lignicolous species with cheiroid, digitate, palmate, and/or dictyosporous conidia. Related with the extant genus Dictyosporium, recorded worldwide from dead wood, decaying leaves, and palm material. |
 Lophiostoma compressum, an extant wood fungus with dictyospores |
|
Sporonites[12] |
|
|
Fungal Spores |
A True Fungus, member of the family Eumycota inside Eumycetes. It is found associated with Wood Cuticles, Pollen and Spores, interpreted as some sort of parasitism. This genus is found mostly associated with the Blanowice Brown Coals, especially on associated Boreholes. |
.jpg.webp) Sporonites was probably related with a plant parasite fungus or a litter fungus, such as Dicranophora |
Invertebrates
Ichnofossils
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Unlined meniscate burrows |
Saltwater/Blackish burrow-like ichnofossils. Taenidium is a meniscate backfill structure, usually considered to be produced by an animal progressing axially through the sediment and depositing alternating packets of differently constituted sediment behind it as it moves forward. |
||
|
|
Small branching burrows |
Saltwater/Blackish burrow-like ichnofossils. Interpreted as the feeding burrow of a sediment-ingesting animal.[15] A more recent study has found that Scoloplos armiger and Heteromastus filiformis, occurring in the German Wadden Sea in the lower parts of tidal flats, make burrows that are homonymous with numerous trace fossils of the ichnogenus.[16] |
 Illustration of Chondrites bollensis | |
|
|
Cylindrical or elliptical curved/tortuous trace fossils |
Saltwater/Blackish burrow-like ichnofossils. Planolites is really common in all types of the Ciechocinek Formation deposits. It is referred to vermiform deposit-feeders, mainly Polychaetes, producing active Fodinichnia. It is controversial, since is considered a strictly a junior synonym of Palaeophycus.[18] |
 Example of Planolites fossil | |
|
Palaeophycus[17] |
|
|
Straight or gently curved tubular burrows. |
Saltwater/Blackish burrow-like ichnofossils. Palaeophycus is less common than Planolites in deposits of the Ciechocinek Formation. On the Kozlowice outcrop however there are numerous specimens occur, interpreted as the result of passive filling of polychaete burrows.[17] |
 Example of Palaeophycus fossil |
|
|
Simple, unbranched, horizontal cylinder traces |
Saltwater/Blackish burrow-like ichnofossils. There are a few specimens in the Kozlowice outcrop. It is interpreted as a grazing trail or Fodinichnia, produced at shallow depth in sediment by Polychaetes and Priapulids.[17] |
 Example of Helminthopsis fossil | |
|
Gyrochorte[17] |
|
|
Winding, horizontal, double ridge burrows, separated by a median groove. |
Saltwater/Blackish burrow-like ichnofossils. On the Kozlowice strata, only a few specimens were observed. Gyrochorte is interpreted as a result of active digging on the sediment by a deposit-feeding worm-like animal, probably Annelid or similar kinds of creatures.[17] Some modern species such as the Polychaete Pectinaria and some Scaphopoda mollusks let similar traces on the sand. Is a genus fond mostly on moderate to moderately high energy environments, including bars, shorefaces of beach complexes, storm-dominated shelves, and embayment areas?[19] |
|
|
Protovirgularia[17] |
|
|
Bilobate trace fossil |
Saltwater/Blackish and Freshwater bottom Trace Fossils. Protovirgularia is a Repichnia form, ascribed to the activity of Bivalves, leaving a trace due to the rhythmic action of a foot.[17] |
|
|
|
Horizontal and subhorizontal, branching tunnels,cylindrical or elliptical in cross-section, displaying elongated striation on exterior of burrow casts |
Saltwater/Blackish burrow-like ichnofossils. On the Kozłowice section a series of sand-filled tunnels of Spongeliomorpha are preserved as Endichnia within mudstone, preserved as Tubular Tempestites (Storm-burrow filling), open tubes produced by animals burrowing in a stable, stiff or firm sub strate.[20] Here the tubes occur in one ho rizon with thin sandy laminae and lenses that represent isolated Starved Ripples carried on the muddy sea-floor by storm-generated Traction power network.[20]Spongeliomorpha is believed to come from the domicile of Crustaceans: Anomuras (Probably Eocarcinoidea) and Decapodas (Probably Glypheidae), created as they dig in a firm, semiconsolidated substrate.[17] Local Spongeliomorpha could point to a transgressional Sea water impulse following a short episode of regression.[17] |
||
|
|
Vertical to oblique, unbranched or branched, elongated to arcuate spreite burrow |
Saltwater/Blackish burrow-like ichnofossils. Is common on the Pomerania Region. The overall morphology and details of the burrows, in comparison with modern analogues and neoichnological experiments, suggest Echiurans (spoon worms) or Holothurians (sea cucumbers) with a combined suspension- and deposit-feeding behaviour as potential producers. |
 Teichichnus burrows | |
|
|
"U"-shaped burrows |
Marine-Mangroove Vertical, U-shaped, single-spreite Burrows; unidirectional or bidirectional spreite, generally continuous, rarely discontinuous.[22] Most Diplocraterion show only protrusive spreit, like the local ones, produced under predominantly erosive conditions where the organism was constantly burrowing deeper into the substrate as sediment was eroded from the top. "U"-shaped burrows, such as Diplocraterion, can be constructed by a wide variety of creatures: Polychaeta annelids (Axiothella, Abarenicola and Scolecolepis), Sipunculans (Sipunculus), Enteropneustans (Balanoglossus) and Echiurans (Urechis).[23] |
 Diplocraterion parallelum diagram | |
Annelida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Dictyothylakos[24] |
|
|
Cocoons |
Freshwater Clitellata Cocoons (Oligochaeta and Hirudinea), identified with palynological residues, and through to be tridimensional nets of probable algal origin.[25] Fragmentary mesh-like networks of happiness threads composed of homogenous translucent material. They show the outer wall (hapsine) construction specific to clitellate annelids and lack an alytine (inner) layer. A disorderly meshwork of the hapsine layer and hapsine fibers of unequal thickness, are diagnostic of the type species Dictyothylakos pesslerae. The cocoons Dictyothylakos pesslerae resemble specially those of modern Leechs, and are common on flooded basin sediments, which implies not only the presence of parasitic leeches, but also the presence of large hosts nearby, as has been confirmed on the case of the Ciechocinek Formation, thanks to the presence of not only dinosaurs but also Dipnoi and other freshwater taxa. |
 Example of leech cocoon .jpeg.webp) Placobdella, example of leech |
Brachiopoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A Saltwater Brachiopodan, member of Discinidae inside Discinida. The classification of the Discinidae is rather treated with confusion, due to the description and the identification of either extinct and extant genera and species. The Shells of this genus are abundant on the German Realm of the formation. |
||
Bivalvia
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A Saltwater Oyster, member of "Posidoniidae" inside Ostreoida. A genus classified under "Posidonia bronni". |
 | |
|
|
Cunchs |
A Saltwater Clam, member of Inoceramidae inside Myalinida. This genus resemble the Modern Pearl Oysters, although it was more likely a clam. The specimens are rather complete. |
||
|
|
Cunchs |
A Saltwater Clam, member of Inoceramidae inside Myalinida. Pseudomytiloides dubius is a possible junior synonym. This genus resembles the Modern Pearl Oysters, although it was more likely a clam. The specimens are rather complete. It is the most common bivalve found locally. |
 Parainoceramya | |
|
|
Cunchs |
A Saltwater Clam, member of Lucinidae inside Lucinida. Very abundant on the layers |
| |
|
|
Cunchs |
A Saltwater Clam, member of Pholadomyidae inside Pholadomyida. Rather common, but less abundant than other local genera |
||
|
|
Cunchs |
A Saltwater Clam, member of Cuspidariidae inside Anomalodesmata. Marginal Marine to Mangrove swamp mollusk fauna, present on a rather large degree of salinities. |
||
|
|
Cunchs |
A saltwater Clam, member of the family Tellinidae inside Cardiida. Thousands of individuals were found associated with the same level. |
.jpg.webp) Example of extant specimen of Macoma | |
|
|
Cunchs |
A saltwater Clam, member of the family Veneridae inside Venerida. A lump was found containing numerous fauna of clams belonging to the genus Cytherea. Is now believed that the liasic forms defined by him as Cytherea they are of completely different kinds. |
||
|
|
Cunchs |
A saltwater Clam, member of the family Veneridae inside Venerida. |
 Example of extant specimen of Callista | |
|
Nuculana[32] |
|
|
Cunchs |
A saltwater Pointed Nut Clam, type member of the family Nuculanidae inside Protobranchia. |
|
|
|
Cunchs |
A Saltwater Scallop, member of Oxytomidae inside Pectinida. Depositional environments in the Polish Basin at this time did not host organisms normally associated with well-oxygenated marine conditions. Meleagrinella substriata is the most common bivalve found on the Toarcian Polish Basin, linked with all the marine Deposits. |
 Meleagrinella | |
|
Pseudomonotis[34] |
|
|
Cunchs |
A Saltwater Scallop, member of Pseudomonotidae inside Pectinida. The deposition of the specimens, on the East shore of the marine Toarcian transgression, suggests a mass mortality event of bivalves. |
|
|
|
Cunchs |
A Saltwater Mussel, member of Mytilidae inside Mytilida. Wyżyna Krakowsko-Częstochowska there is abundant the microfauna represented by foraminifers along with also mussels, unfortunately, due to the poor state of preservation not determinate (Probably Modiolus sp.). |
 Example of extant specimen of Modiolus | |
|
|
Cunchs |
A Freshwater Mussel, type member of the family Unionoida inside Palaeoheterodonta. The Only major Freshwater Bivalve recovered on the Formation. |
 Example of extant specimen of Unio | |

Gastropoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A holoplanktonic Snail, type member of the family Coelodiscidae inside Prosobranchia. This is the oldest known holoplanktonic gastropod, thanks to bilateral symmetrical shells as an adaption to active swimming. Also the most common of the sea snails on the Clay Pit of Dobbertin, where is one of the most varied in size terms, with some of the biggest specimens of this snail from the Lower Toarcian know.[36] It has been related to large floating driftwood as one of the primary settlers.[36] |
||
|
|
Cunchs |
A Sea Snail, member of Cerithiinae inside Caenogastropoda. The main recovered gasteropod from the Green Series. Extant genus. |
| |
|
|
Cunchs |
A Sea Snail, member of Procerithiidae inside Caenogastropoda. |
||
|
Rhynchocerithium[37] |
|
|
Cunchs |
A Sea Snail, member of Procerithiidae inside Caenogastropoda. The local assigned Francocerithium? sp and Francocerithium kochi from the older Pliensbachian strat can be all Rhynchocerithium sp. |
|
|
Ptychomphalus[37] |
|
|
Cunchs |
A Sea Snail, member of Eotomariidae inside Pleurotomarioidea. The species may be identical to the questionable "Ptychomphalus" theodorii. |
|
|
|
Cunchs |
A Sea Snail, member of Settsassiidae inside Hypsogastropoda. Turriculate, slender shells. |
||
|
Maturifusus[37] |
|
|
Cunchs |
A Sea Snail, type member of Maturifusidae inside Hypsogastropoda. Limited to the Grimmen Clay Pit |
|
|
Cylindrobullina[37] |
|
|
Cunchs |
A Sea Snail, type member of Cylindrobullinidae inside Architectibranchia. Diversified through the Polish realm, but more common on Grimmen. |
|
|
|
Cunchs |
A Sea Snail, member of Acteoninidae inside Prosobranchia. The most diverse on the Polish realm, also the most common. |
||
|
|
Cunchs |
A sea Snail, member of Zygopleuridae inside Murchisoniina. More abundant on the German Realm |
||
|
|
Cunchs |
An opisthobranch sea Snail, member of Murchisonellidae inside Pyramidelloidea. Trend to be limited to northern deposits |
||
|
Kalchreuthia[37] |
|
|
Cunchs |
A minute sea Snail, member of Iravadiidae inside Truncatelloidea. This Genus is abundant on Pomerania, but trends to be absent on mostly of the Grimmen Pit strata. |
|
|
Tricarilda[37] |
|
|
Cunchs |
A minute sea Snail, member of Mathildidae inside Allogastropoda. The Tricarilda? sp. of Grimmen is maybe the same as the assigned specimens Tricarilda? sp. of Reinberg |
|
|
Lewisiella[37] |
|
|
Cunchs |
A Sea false top Snail, member of Ataphridae inside Trochoidea. Lewisiella nuda is also known from Franconia, with only 10 specimens from Grimmen. |
|
|
|
Cunchs |
A river or "mystery" Snail, type member of the family Viviparidae inside Viviparoidea. In the area of the northern and north-western edge of the Swiętokrzyskie Mountains, the sea fauna has been found sporadically, but there are several finds of freshwater forms. Viviparus main identified snail on the Formation, and a typical form associated with Deltaic-Fluvial Deposits. The major Toarcian concentration of this genus is found on Gorzków. |
 Example of extant specimen of Viviparus | |

Cephalopoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Shells |
An Ammonite, member of the family Hildoceratidae inside Ammonoidea. |
 Eleganticeras | |
|
|
Shells |
An Ammonite, member of the family Hildoceratidae inside Ammonoidea. |
| |
|
|
Shells |
An Ammonite, member of the family Hildoceratidae inside Ammonoidea. |
| |
|
|
Shells |
An Ammonite, member of the family Phylloceratidae inside Ammonoidea. Among the largest ammonites found on the Green Series, with specimens over 60 cm |
| |
|
|
Shells |
An Ammonite, member of the family Lytoceratidae inside Ammonoidea. |
| |
|
|
Shells |
An Ammonite, member of the family Lytoceratidae inside Ammonoidea. |
| |
|
|
Shells |
An Ammonite, member of the family Harpoceratinae inside Ammonoidea. |
 Pseudolioceras | |
|
|
Shells |
An Ammonite, type member of the family Dactylioceratinae inside Ammonoidea. Most common ammonite found on the Green Series and the different erratic boulders, as is the most common on the German realm, on the north and the south, with several specimens of different sizes. |
| |
|
|
Shells |
An Ammonite, member of the family Dactylioceratinae inside Ammonoidea. |
||
|
|
Multiple Specimens. |
A Belemnoidean. member of the family Passaloteuthididae inside Belemnitida. |
||
|
|
Multiple Specimens. |
A Belemnoidean. member of the family Megateuthididae inside Belemnitida. |
||
|
|
Multiple Specimens. |
A Belemnoidean. member of the family Megateuthididae inside Belemnitida. |
| |
|
|
Multiple Specimens. |
A Mesoteuthoidean, member of the family Beloteuthidae. Is a relatively small genus. |
.jpg.webp) Beloteuthis | |
|
|
Multiple Specimens. |
A Vampire Squid, member of Teudopsidae inside Vampyropodan. Related to the modern Vampyroteuthis infernalis. |
| |
|
|
Multiple Specimens. |
A Loligosepiidae Loligosepiidan (Vampyromorpha). Related to the modern Vampyroteuthis infernalis. Gladii of Loligosepia can be distinguished from Jeletzkyteuthis by the transition lateral field/hyperbolar zone. Described originally as Belopeltis bollensis. |
||








Echinodermata
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A Brittle Star, member of Ophiuroidea inside Echinodermata. |
||
|
|
|
A Brittle Star, member of Ophiuroidea inside Echinodermata. It is relatively less abundant than in coeval strata. |
 Palaeocoma | |
|
|
|
A Brittle Star, member of Ophiodermatidae inside Echinodermata. Still alive today. On the Green Series, is related to deep basinal deposits. |
| |
|
|
|
A Sea Star, member of Astropectinidae inside Asteroidea. Related to freshwater debris, that probably caused changes to salinity and mass mortality of Echinoderms. |
 Archastropecten | |
|
|
|
A sea lily, member of Isocrinidae inside Crinoidea. |
||
|
|
Stems |
A sea lily, member of Pentacrinitidae inside Isocrinida. A pelagic Sea lilly, related with floating logs. |
| |
|
|
Spines |
A Sea Urchin, member of Echinoidea inside Echinodermata. |
||
|
|
|
A sea Cucumber, member of Holothuriida inside Holothuroidea. Sea cucumbers from marginal marine settings. |
||
|
|
|
A pelagic sea Cucumber, member of Elasipodida inside Holothuroidea. Open pelagic settings sea cucumbers probably washed to nearshore deposits |
||
|
|
|
A sea cucumber, member of the family Synaptidae inside Apodida. Due o the similarity of the fossils of this genus with the fishing hooks of some extant Synaptidae (exmpl. Chirodota japonica) is assumed that Ancistrum is a synaptid holothurian. |
||
|
|
|
A sea Cucumber, probably a member of the family Synaptidae inside Apodida. The systematic position of the fossil is uncertain. It has been seen an "Ankerplatten" on the holotype, that is derived from Synaptids. |
||
|
|
|
A sea Cucumber, type member of the family Chiridotidae inside Apodida. It has an edge of the wheel rim with numerous sharp teeth. Also a conical shape on the outside of the wheel. The "cogs" of Chirodota - like on the genus Myriotrochus- precisely taken are very flat, round cups that are so in the skin of the animal is oriented so that its concavity points towards its surface. |
 Extant Chirodota | |
|
|
|
A sea Cucumber, member of the family Dendrochrotae inside Dendrochirotida. It isn't comparable to none of the modern sea Cucumbers. It has, however, lamellas like the ones found on species of the genus Echinocucumis, and also the genus Cucumaria. |
.jpg.webp) Extant Cucumaria, Crux may have been similar in appearance | |


Crustacea
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Chelae |
A marine Hermit Crab, member of the family Paguridae inside Decapoda. It has been found on deposits with marginal marine to Mangroove influence. |
||
|
|
Various specimens. |
A Marine Decapodan, probably a member of Eocarcinoidea inside Anomura. It is among the oldest found Crabs worldwide. The genus is known locally mostly due to fragmentary specimens, related to less saline conditions. |
||
|
|
Various specimens. |
A Marine Decapodan, type member of the family Glypheidae inside Decapoda. The most complete Crustacean found on the formation |
| |
|
Proeryon[55] |
|
|
Single specimen with preserved upper thorax and chelae |
A Marine Decapodan, member of the family Proeryoninae inside Polychelida. Resemble Proeryon hartmani show less adaptations to hunt for small nectobenthic preys than other relatives, being abundant on Oyster-filled waters. There is a relative abundance of the genus in deep-water settings from the Toarcian onward.[56] |
 Proeryon |
|
|
Cunchs |
A Freshwater Clam shrimp (Phyllopodan) of the family Lioestheriidae. They fed on detritus, being very small slow moving, nektonic organisms that filter fed as they floated. The specimens are numerous here and form thin layers, being the most abundant invertebrate recovered on all the formation. The presence of Palaeestheria marks the appearance of less saline conditions, as this is a mostly freshwater genus. The local Phyllopods are related with a great amount of freshwater debris (especially plants), and suggest seasonal changes on the rivers on the Toarcian Polish Basin. On Kozłowice, there is an association of the ichnogenus Planolites and phyllopods of this genus, which reflects a gradual decline in water salinity during a sea level high standard regression. |
 Palaeestheria | |
|
|
Valves |
A Marine Ostracodan of the family Pontocyprididae. Small marine ostracods related with abundant Green Algae environments |
||
|
Ambigocythere[58] |
|
|
Valves |
A Marine Ostracodan with incertade Sedis assignation. The specimens of this genus are rather fragmentary. |
|
|
Infracytheropteron[59] |
|
|
Valves |
A Marine Ostracodan of the family Protostomia. The specimens of this genus are rather fragmentary and of uncertain nature. |
|
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. This genus is the main reported on the marine facies of the Dobbertin Clay Pit. |
||
|
Ogmoconcha[63] |
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. Is probably present on all the clay pits, although the other locations specimens were not published. |
|
|
Ledahia[64] |
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. One of the genera that reflect better the migration patterns of ostracodans on the Pliensbachian-Toarcian boundary. |
|
|
Pseudohealdia[64] |
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. The genus is rare on the layers. |
|
|
Hermiella[61] |
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. This genus is the main reported on the marine facies of the Dobbertin Clay Pit. |
|
|
Cytheropteron[65] |
|
|
Valves |
A Marine Ostracodan of the family Cytheruridae inside Podocopida. Is rare and the specimens found are rather incomplete. |
|
|
Nanacythere[65] |
|
|
Valves |
A Marine Ostracodan of the family Cytheruridae inside Podocopida. The most abundant genus on the Grimmen Clay Pit. |
|
|
Procytherura[66] |
|
|
Valves |
A Marine Ostracodan of the family Cytheruridae inside Podocopida. A genus with well preserved specimens locally. |
|
|
|
Valves |
A Marine Ostracodan of the family Protocytheridae inside Podocopida. Common, and associated with benthonic deposits. This genus maybe was able to resist relative changes in salinity. |
||
|
Kinkelinella[58] |
|
|
Valves |
A Marine Ostracodan of the family Protocytheridae inside Podocopida. A genus related with fish fossils and anoxic bottoms. |
|
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. Abundant and diverse, is found associated with Ammonite shells. |
||
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. Less abundant than the genus Bairdia, is present on layers where wood debris is more abundant. |
||
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. Know only from the Green Series, is a rather unusual and complex genus. |
||
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. It is a relatively abundant genus, but based on incomplete material. |
||
|
Polycope[59] |
|
|
Valves |
A Marine Ostracodan of the family Polycopidae inside Cladocopina. Scarce but well preserved specimens. |
|

Arachnida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Single Incomplete Specimen. |
A spider, possible member of the superfamily Palpimanoidea.[71] It is the first confirmed spider from the lower Jurassic, and a rare find, probably washed to the sea due to a hard wind related to hurricane action, present due to the measured monsoonal conditions of the formation, as on the rest of the Lower-Middle Toarcian strata. Probably a ground-dwelling predator that hunted the abundant insect fauna present on the layers.[71] With a robust and well-armed legs I, directed forwards give the suggestion that they were preycapture appendages, a morphology typical of a sit-and-wait predator, while the short legs III are more typical on web spiders, especially Orbweavers, but also found on Palpimanoids, but not on that that are common substrate dwellers, that had legs more equal.[71] Seppo was probably not a habitual ground dweller, with armoured front legs related to capturing dangerous prey, such as many palpimanoids today are Araneophagous, for example.[71] |
 Seppo koponeni reconstruction on a Tree, as suggested for its Leg III morphology | |
Insecta
Insects are a common terrestrial animals that were probably drifted to the sea due to Moonsonal conditions present on the Ciechocinek Formation.[72]
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Wing Scales |
A Moth, member of the family Eolepidopterigidae inside Lepidoptera. They are essential fossils for the Development of the color on Lepidopterans. |
||
|
Zalmonites[74] |
|
|
Specimens |
A Grasshopper, Incertade Sedis inside Orthoptera. |
|
|
|
Specimens |
A Grasshopper, member of the family Elcanidae inside Orthoptera. The species P. magna is among the largest Orthopterans of the Jurassic, while P. minima is among the smallest. |
||
|
|
Specimens |
A Grasshopper, member of the family Elcanidae inside Orthoptera. |
||
|
|
Specimens |
A Grasshopper, member of the family Elcanidae inside Orthoptera. |
||
|
|
Specimens |
A Grasshopper, type member of the family Locustopsidae inside Orthoptera. |
||
|
|
Specimens |
A Grasshopper, member of the family Locustopsidae inside Orthoptera. |
||
|
|
Specimens |
A Grasshopper, type member of the family Locustopsidae inside Orthoptera. |
||
|
|
Specimens |
A Locust, type member of the family Locustidae inside Orthoptera. |
||
|
|
Specimens |
A Grasshopper, member of the family Regiatidae inside Orthoptera. |
||
|
|
Specimens |
A Cricket-like Grasshopper, type member of the family Protogryllidae inside Orthoptera. |
||
|
Griphopteron[80] |
|
|
Specimens |
A Cricket-like insect, member of the family Blattogryllidae inside Eoblattida. |
|
|
|
Specimens |
A Stick Insect, member of the family Aerophasmidae inside Phasmatodea. |
||
|
Aenne[81] |
|
|
Specimens |
The oldest known non biting Midge (Chironomidae) inside Diptera |
|
|
Oryctochlus[81] |
|
|
Specimens |
The oldest known non biting Midge (Chironomidae) inside Diptera |
|
|
Podonomius[81] |
|
|
Specimens |
The oldest known non biting Midge (Chironomidae) inside Diptera |
|
|
Dobbertiniopteryx[82] |
|
|
Specimens |
A small winter Stonefly, member of the family Capniidae inside Plecoptera. |
|
|
|
Specimens |
A Crane Fly of the family Limoniidae inside Diptera. |
||
|
|
Specimens |
A Crane Fly of the family Limoniidae inside Diptera. A. nana can be the smallest Crane fly of the Jurassic. |
||
|
Eotipula[74] |
|
|
Specimens |
A Crane Fly of the family Limoniidae inside Diptera. |
|
|
Phryganidium[74] |
|
|
Specimens |
A Crane Fly, member of the family Limoniidae inside Diptera. |
|
|
Grimmenia[83] |
|
|
Specimens |
A Crane Fly of the family Limoniidae inside Diptera. |
|
|
Rhaetomyia[80] |
|
|
Specimens |
A Phantom midge of the family Chaoboridae inside Diptera. |
|
|
Grimmyia[80] |
|
|
Specimens |
A Snipe Fly, member of the family Rhagionidae inside Diptera. |
|
|
Liassobrachyceron[85] |
|
|
Specimens |
A Snipe Fly, member of the family Rhagionidae inside Diptera. |
|
|
Palaeobrachyceron[80] |
|
|
Specimens |
A Snipe Fly, member of the family Rhagionidae inside Diptera. |
|
|
Antefungivora[80] |
|
|
Specimens |
A Fly, type member of the family Antefungivoridae inside Diptera. |
|
|
Archibio[80] |
|
|
Specimens |
A Fly, member of the family Antefungivoridae inside Diptera. |
|
|
Pleciofungivora[80] |
|
|
Specimens |
A Fly, member of the family Pleciofungivoridae inside Diptera. |
|
|
Archirhyphus[80] |
|
|
Specimens |
||
|
Protorhyphus[77] |
|
|
Specimens |
A Fly, type member of the family Protorhyphidae inside Diptera |
|
|
Protobrachyceron[86] |
|
|
Specimens |
A Fly, member of the family Protobrachyceridae inside Diptera |
|
|
Heterorhyphus[80] |
|
|
Specimens |
A Fly, type member of the family Heterorhyphidae inside Diptera |
|
|
Eoditomyia[80] |
|
|
Specimens |
||
|
Archipleciomima[80] |
|
|
Specimens |
||
|
Protoplecia[80] |
|
|
Specimens |
||
|
Mailotrichocera[87] |
|
|
Specimens |
A Winter Crane Fly, member of the family Trichoceridae inside Diptera. |
|
|
Nannotanyderus[88] |
|
|
Specimens |
A primitive Crane Fly, member of the family Tanyderidae inside Diptera. |
|
|
Praemacrochile[88] |
|
|
Specimens |
A primitive Crane Fly, member of the family Tanyderidae inside Diptera. |
|
|
|
Specimens |
A Phantom crane fly, member of the family Ptychopteridae inside Diptera. |
||
|
|
Specimens |
A Phantom crane fly, member of the family Ptychopteridae inside Diptera. |
||
|
|
Specimens |
A Phantom crane fly, member of the family Ptychopteridae inside Diptera. |
||
|
Tanypsycha[88] |
|
|
Specimens |
A Moth fly, member of the family Psychodidae inside Diptera. |
|
|
Liassopsychodina[88] |
|
|
Specimens |
A Moth fly, member of the family Psychodidae inside Diptera. |
|
|
Mesorhyphus[86] |
|
|
Specimens |
A Wood Gnat, member of the family Anisopodidae inside Diptera. |
|
|
Metatrichopteridium[90] |
|
|
Specimens |
A rare Fly, member of the family Hennigmatidae inside Diptera. |
|
|
Orthophlebia[77] |
|
|
Specimens |
A hangingfly, type member of the family Orthophlebiidae inside Mecoptera. |
|
|
Mesopanorpa[77] |
|
|
Specimens |
A hangingfly, type member of the family Orthophlebiidae inside Mecoptera. |
|
|
Neorthophlebia[77] |
|
|
Specimens |
A hangingfly, member of the family Bittacidae inside Mecoptera. |
|
|
Parabittacus[91] |
|
|
Specimens |
A hangingfly, member of the family Bittacidae inside Mecoptera. |
|
|
Mesobittacus[77] |
|
|
Specimens |
A hangingfly, member of the family Bittacidae inside Mecoptera. |
|
|
Pseudopolycentropus[74] |
|
|
Specimens |
A scorpionfly, type member of the family Pseudopolycentropodidae inside Mecoptera. |
|
|
Mesochorista[74] |
|
|
Specimens |
A Scorpionfly, member of the family Permochoristidae inside Mecoptera. |
|
|
Liasocoris[92] |
|
|
Specimens |
A True bug, Incertade sedis inside Hemiptera. |
|
|
|
Specimens |
A Planthopper, the type genus of the family Fulgoridiidae inside Hemiptera. The colossal abundance of the genus maybe it's related to a preference for seashore habitats. Some specimens are indentinguible, making possible some species synonymous. |
||
|
Metafulgoridium[77] |
|
|
Specimens |
A Planthopper, member of the family Fulgoridiidae inside Hemiptera. |
|
|
Margaroptilon[77] |
|
|
Specimens |
A Planthopper, member of the family Fulgoridiidae inside Hemiptera. |
|
|
Cixiites[77] |
|
|
Specimens |
A Planthopper, member of the family Fulgoridiidae inside Hemiptera. |
|
|
Procercopis[77] |
|
|
Specimens |
A Froghopper, type member of the family Procercopidae inside Hemiptera. |
|
|
Liadopsylla[93] |
|
|
Specimens |
A jumping plant louse, member of the family Liadopsyllidae inside Hemiptera. |
|
|
Hadrocoris[86] |
|
|
Specimens |
A true Bug, member of the family Hadrocoridae inside Hemiptera. |
|
|
Acromocoris[94] |
|
|
Specimens |
A water boatman, member of the family Corixidae inside Hemiptera. |
|
|
Progonocimex[86] |
|
|
Specimens |
A Moss bug, type member of the family Progonocimicidae inside Hemiptera. |
|
|
Eocercopis[77] |
|
|
Specimens |
A Moss bug, member of the family Progonocimicidae inside Hemiptera. |
|
|
Archicercopis[77] |
|
|
Specimens |
A Moss bug, member of the family Progonocimicidae inside Hemiptera. |
|
|
|
Specimens |
A Coleorrhynchan, member of the family Probascanionidae inside Hemiptera. |
||
|
Archegocimex[74] |
|
|
Specimens |
A shore bug, member of the family Archegocimicidae inside Hemiptera. |
|
|
Progonocoris[86] |
|
|
Specimens |
A shore bug, member of the family Archegocimicidae inside Hemiptera. |
|
|
Anosmus[77] |
|
|
Specimens |
A shore bug, member of the family Archegocimicidae inside Hemiptera. |
|
|
Diatillus[93] |
|
|
Specimens |
A shore bug, member of the family Archegocimicidae inside Hemiptera. |
|
|
Pachymeridium[13] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Sisyrocoris[93] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Hypocimex[77] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Apsicoria[77] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Cathalus[77] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Psychrocoris[86] |
|
|
Specimens |
A true Bug, member of the family Pachymeridiidae inside Hemiptera. |
|
|
Cuneocoris[86] |
|
|
Specimens |
A true Bug, member of the family Cuneocoridae inside Hemiptera. |
|
|
Apopnus[93] |
|
|
Specimens |
A creeping water bug, member of the family Naucoridae inside Hemiptera. |
|
|
Aphlebocoris[74] |
|
|
Specimens |
A creeping water bug, member of the family Naucoridae inside Hemiptera. |
|
|
|
Specimens |
A leafhopper, type member of the family Archijassidae inside Hemiptera. |
||
|
|
Specimens |
A leafhopper, type member of the family Archijassidae inside Hemiptera. |
||
|
|
Specimens |
A leafhopper, member of the family Archijassidae inside Hemiptera. |
||
|
|
Specimens |
A cicadomorphan, member of the family Hylicellidae inside Hemiptera. |
||
|
|
Specimens |
A Sternorrhynchan, member of the family Protopsyllidiidae inside Hemiptera. |
||
|
Archiconiopteryx[96] |
|
|
Specimens |
A Stemorrhynchan, type member of the family Archiconiopterygidae inside Hemiptera. |
|
|
|
Specimens |
A Sternorrhynchan, member of the family Protopsyllidiidae inside Hemiptera. |
||
|
|
Specimens |
An aphid, member of the family Oviparosiphidae inside Hemiptera. |
||
|
Liassochrysa[97] |
|
|
Specimens |
A Neuropteran of the family Mantispidae. It is the earliest know Chrysopid |
|
|
Prohemerobius[98] |
|
|
Specimens |
A lacewing, type member of the family Prohemerobiidae inside Neuroptera. |
|
|
|
Specimens |
A lacewing, type member of the family Prohemerobiidae inside Neuroptera. |
||
|
Mesosmylina[99] |
|
|
Specimens |
A lance lacewing, member of the family Osmylidae inside Neuroptera. |
|
|
Polyosmylus[80] |
|
|
Specimens |
A lacewing, Incertade sedis inside Neuroptera. |
|
|
Mesoleon[80] |
|
|
Specimens |
A lacewing, Incertade sedis inside Neuroptera. |
|
|
|
Specimens |
A lacewing, Incertade sedis inside Neuroptera. |
||
|
|
Specimens |
A lacewing, Incertade sedis inside Neuroptera. |
||
|
Melamnous[77] |
|
|
Specimens |
A lacewing, Incertade sedis inside Neuroptera. |
|
|
Dobbertinia[77] |
|
|
Specimens |
An alderfly, member of the family Sialidae inside Megaloptera. |
|
|
Mesotrichopteridium[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Metarchitaulius[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Parataulius[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Nannotrichopteron[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Trichopteridium[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Necrotaulius[80] |
|
|
Specimens |
A caddisfly, type member of the family Necrotauliidae inside Trichoptera. |
|
|
Epididontus[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Archiptilia[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Paratrichopteridium[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Pararchitaulius[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Palaeotaulius[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
Pseudorthophlebia[80] |
|
|
Specimens |
A caddisfly, member of the family Necrotauliidae inside Trichoptera. |
|
|
|
Specimens |
A finger-net caddisfly, member of the family Philopotamidae inside Trichoptera. |
||
|
Solenoptilon[77] |
|
|
Specimens |
A lacewing, type member of the Family Solenoptilidae inside Neuroptera. |
|
|
Epigambria[77] |
|
|
Specimens |
A lacewing, member of the Family Epigambriidae inside Neuroptera. |
|
|
Apeirophlebia[77] |
|
|
Specimens |
A silky lacewing, member of the Family Psychopsidae inside Neuroptera. An unexpected giant Silky lacewing, with a size up to 7 cm. |
|
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. Very large dragonfly, with a wingspan of 13 cm |
||
|
Liadothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Petrothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Parelthothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Anomothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Rhabdothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Temnostigma[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Pycnothemis[77] |
|
|
Specimens |
A Dragonfly, Incertade sedis inside Odonata. |
|
|
Archithemis[77] |
|
|
Specimens |
A Dragonfly, member of the family Archithemistidae inside Odonata. |
|
|
Heterothemis[102] |
|
|
Specimens |
A Dragonfly, member of the family Liassogomphidae inside Odonata. |
|
|
Anisozygopteron[77] |
|
|
Specimens |
A Dragonfly, member of the family Myopophlebiidae inside Odonata. |
|
|
Selenothemis[77] |
|
|
Specimens |
A Dragonfly, type member of the family Selenothemistidae inside Odonata. |
|
|
Dialothemis[103] |
|
|
Specimens |
A Dragonfly, member of the family Selenothemistidae inside Odonata. |
|
|
Eosagrion[86] |
|
|
Specimens |
A Dragonfly, member of the family Eosagrionidae inside Odonata. |
|
|
Turanopteron[80] |
|
|
Specimens |
A Dragonfly, member of the family Asiopteridae inside Odonata. "Sphenophlebia pommerana" is a junior synonym. |
|
|
Grimmenopteron[80] |
|
|
Specimens |
A Dragonfly, member of the family Liassophlebiidae inside Odonata. |
|
|
Trigonophlebia[80] |
|
|
Specimens |
A Dragonfly, member of the family Heterophlebiidae inside Odonata. |
|
|
Heterophlebia[80] |
|
|
Specimens |
A Dragonfly, member of the family Heterophlebiidae inside Odonata. |
|
|
Grimmaratavites[104] |
|
|
Specimens |
A Wood Wasp, member of the family Karatavitidae inside Hymenoptera. |
|
|
Liadobracona[105] |
|
|
Specimens |
A Wasp, member of the family Ephialtitidae inside Hymenoptera. |
|
|
|
Specimens |
A Wasp, member of the family Ephialtitidae inside Hymenoptera. |
||
|
|
Specimens |
A stem Sawfly, member of the family Sepulcidae inside Hymenoptera. |
||
|
Mesoblattina[107] |
|
|
Specimens |
A Cockroach, type member of the family Mesoblattinidae inside Blattodea. |
|
|
|
Specimens |
A Cockroach, type member of the family Caloblattinidae inside Blattodea. |
||
|
|
Specimens |
A Cockroach, type member of the family Raphidiomimidae inside Blattodea. |
||
|
|
Specimens |
A Cockroach, member of the family Blattulidae inside Dicondylia. |
||
|
|
Specimens |
A Cockroach, type member of the family Blattulidae inside Dicondylia. Member of an extinct cockroach-like family which was widely distributed around the world and lasted from Late Triassic to Cretaceous. |
||
|
Dicronemoura[80] |
|
|
Specimens |
A stonefly, member of the family Perlariopseidae inside Plecoptera. |
|
|
Liassopsocus[80] |
|
|
Specimens |
A Paraneopteran, member of the family Psocidiidae inside Permopsocida. |
|
|
Archipsylla[80] |
|
|
Specimens |
A Paraneopteran, member of the family Archipsyllidae inside Permopsocida. |
|
|
Undacypha[80] |
|
|
Specimens |
A Primitive Winged insect, Dicondylian of uncertain Placement. |
|
|
Liadoptilia[80] |
|
|
Specimens |
A Primitive Winged insect, Dicondylian of uncertain Placement. |
|
|
Trichorthophlebia[80] |
|
|
Specimens |
A Primitive Winged insect, Dicondylian of uncertain placement. |
|
|
Obotritagrion[80] |
|
|
Specimens |
A Primitive Winged Insect, member of the family Protomyrmeleontidae inside Dicondylia. |
|
|
Protomyrmeleon[80] |
|
|
Specimens |
A Primitive Winged Insect, member of the family Protomyrmeleontidae inside Dicondylia. "Zirzipanagrion quadriordinum" is a junior synonym. |
|
|
Parnidium[74] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Polypamon[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Bathygerus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Plastonebria[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Plastobuprestites[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Nannoodes[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Pseudocyphon[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Keleusticus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Allognosis[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Enamma[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Pseudoprionites[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Nebrioides[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Paracurculium[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Apioderes[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Bareus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Anypostatus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Periboloptera[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Clinomerus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Anomerus[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Masselytron[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Oxytoroptera[110] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Ecthlimma[77] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Onkedodimus[77] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Apsychus[77] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
||
|
Anancaeon[77] |
|
|
Specimens |
A Beetle, Incertade sedis inside Coleoptera. |
|
|
Hydrobiites[77] |
|
|
Specimens |
A Beetle, member of the family Permosynidae inside Coleoptera. |
|
|
Thoracotes[74] |
|
|
Specimens |
A bark gnawing beetle, member of the family Trogossitidae inside Coleoptera. |
|
|
Eurynucha[74] |
|
|
Specimens |
A jewel beetle, member of the family Buprestidae inside Coleoptera. |
|
|
Coptogyrinus[74] |
|
|
Specimens |
A whirligig beetle, member of the family Gyrinidae inside Coleoptera. |
|
|
Carabites[74] |
|
|
Specimens |
A ground beetle, member of the family Carabidae inside Coleoptera. |
|
|
Nele[80] |
|
|
Specimens |
An ice crawler, member of the family Bajanzhargalanidae inside Grylloblattodea. One of the main Floor insects found. |
|
|
Prosepididontus[80] |
|
|
Specimens |
A winged insect, member of the family Geinitziidae inside Reculida. |
|
|
Geinitzia[80] |
|
|
Specimens |
A winged insect, member of the family Geinitziidae inside Reculida. |
|
Vertebrates
Actinopteri
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Freshwater/Brackish/Marine Osteichthyes, incertade sedis inside Teleostei. Inconclusive remains of fish (mostly teeth) are found mostly on Częstochowa-Wieluń region. The Remains of fish are rather rare on the Polish Realm of the Formation, and found related with the Silesian coast, where embayments and lagoons were formed. The remains were not compared. The lack of typical marine fauna indicates that the Toarcian sea transgression did not cause the development of normal-marine conditions in the Polish Basin. "Type A" and "Type B" includes fragmentary remains that resemble the genera Dapedium and andya, with rounded teeth. "Type C" is more similar to the genus Ptycholepis. Finally, "Type D" & "Type E" resemble the Amiiformes and Osteoglossiformes. |
||
|
Pteroniscus[1] |
|
|
|
A marine/brackish Osteichthyes, member of Palaeonisciformes inside Chondrostei. Fish tooth remains of this genus are found mostly in Kujawy (Aleksandrów I), along with marine foraminifers of the genus Haplophragmoides sp., which indicates a marine deposition. It represents a relict genus and resembles the Late Jurassic Pteroniscus turkestanensis |
|
|
|
|
A marine/brackish Osteichthyes, the youngest representative of the family Saurichthyidae inside Chondrostei. Distinctive for its large jaws, similar to modern Belonidae, it is a genus known from several locations across Europe on the Lower-Middle Jurassic. |
| |
|
|
|
A Freshwater/brackish/marine Osteichthyes, type member of the family Saurichthyidae inside Chondrostei. This genus is known mostly from the late Triassic deposits on Europe, making its appearance on Toarcian strata superficially dubious. It can be a member of the genus Saurorhynchus, although this last one has never been found on Brackish-Prodeltaic Strata. |
| |
|
|
GPIH 4864, Hyomandibula |
A marine Osteichthyes, member of the Chondrosteidae inside Acipenseriformes. This find, which probably originates from the western Baltic basin between Bornholm Island (Denmark) and northeastern Germany, markedly expands the known range of this chondrosteid taxon across the northern part of the strait connecting the Boreal Sea with the Tethys Ocean during the Early Jurassic.[113] The relatively small size of the hyomandibula compared to the largest English material (ca. 40% smaller) may be related to a younger ontogenetic stage of the individual.[113] |
| |
|
Grimmenichthys[114] |
|
|
Articulated, but incompletely preserved specimen |
A marine Osteichthyes, member of the family Pholidophoriformes inside Teleostei. Found Exclusively on the Green Series, this genus was a small ‘pholidophoriform’ teleost. The total length remains unknown due to poor preservation, but probably does not exceed 100 mm. |
|
|
|
|
A marine Osteichthyes, member of the family Pholidophoriformes inside Teleostei. Resmble Pholidophoroides crenulata and P. limbata. |
||
|
|
|
A marine Osteichthyes, member of the family Pholidophoriformes inside Teleostei. |
| |
|
|
Unknown, only cited. |
A marine Osteichthyes, member of the family Pachycormiformes inside Teleostei. The first find of the group in the region. |
||
|
|
|
A marine Osteichthyes, type member of the family Leptolepidae inside Teleostei. |
| |
|
Indeterminate |
|
|
A marine Osteichthyes, member of the family Leptolepidae inside Teleostei. |
||
|
Proleptolepis sp. |
|
|
A marine Osteichthyes, member of the family Leptolepidae inside Teleostei. Proleptolepis in the lower Toarcian of Grimmen suggests that this genus might have had a much wider palaeobiogeographical and temporal distribution, since specimens attributed to Proleptolepis have previously been reported only from the Sinemurian of western Europe. |
||
|
|
Incomplete specimen |
A marine Osteichthyes, member of the family Semionotidae inside Semionotiformes. Dapedium and Tetragonolepis may have used alternative strategies in order to partition food resources efficiently and avoid direct competition. |
| |
|
|
Various specimens |
A marine Osteichthyes, member of the family Semionotidae inside Neopterygii. Stomach content is also preserved on a specimen from Dobbertin, and is composed by arthropod cuticles.[120] |
| |
|
|
Almost complete left prearticular with dentition |
A marine Osteichthyes, Member of the family Pycnodontiformes inside Neopterygii. Is of comparatively small size, suggesting a hypothetical small standard length of the fish of about 7–10 cm at the time of death.[121] It is a taxon that shows clear adaptations to durophagy, inferring a predominant diet of especially hard food items.[121] |
||
|
|
Incomplete skull |
A marine Osteichthyes, type member of the Dapediidae inside Neopterygii.[112] |
| |
|
|
GG 439/4-7 scales |
The oldest confirmed true Gar, member of the Lepisosteidae inside Lepisosteiformes. The oldest reliable lepisosteiform remains. |
 Lepisosteus extant example of Gar | |








Chondrichthyes
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A marine Shark, member of the Hybodontiformes. Related to Hybodus hauffianus and other genera from the south of Germany |
| |
|
|
|
Marine/Brackish/Freshwater Sharks, incertade sedis inside Selachimorpha. Related with the local marine Transgression, the fossils are recovered mostly from an embayment-derived environment. It superficially resembles some Hybodontiformes members (specially Acrodontidae). Together with the rather poor fauna recovered on the main part of the region, the transgression spread out over Mudflats and alluvial Floodplains, to which small amounts of sand were still sporadically supplied during stronger floods. |
||

Sarcopterygii
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ceratodus sp. |
|
|
A freshwater Lungfish, type member of the Ceratodontidae inside Ceratodontiformes. Can be related with Polish Triassic genus Ceratodus silesiacus (Roemer, 1870), that has been known by several isolated tooth plates have been found in the lacustrine Triassic horizon of Krasiejów. Several Impressions of scales found in other Boreholes can belong also to Lugfish. The Tooth plates were found at +819 m depth on Pomerania, on a layer with gray-green mudstone, lenticular lamination and siderite concretions. The abundance of Megaspores indicate a Deltaic or Brackish Environment. |
 Ceratodus reconstruction | |
Ichthyosaurs
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Posterior left half of the cranium. |
An Icthyosaur of the family Stenopterygiidae inside Thunnosauria. A common Toarcian Ichthyosaur, present on multiple layers. The rather exquisite level of preservation has led to know even the coloration. |
 Restoration | |
|
|
Four articulated tail vertebrae. |
An indeterminate ichthyosaur, has been assigned to the species Stenopterygius longifrons |
||
|
|
Partial skull and associated postcranial elements preserved in a concretion |
An indeterminate ichthyosaur. It has an expanded basipterygoid process on the basisphenoid, only currently known in members of the Ophthalmosauridae |
||
|
|
|
An indeterminate ichthyosaur. |
||
|
|
|
An Icthyosaur, type member of Temnodontosauridae inside Neoichthyosauria. Assigned to Ichthyosaurus sp., but also suggested affinities to "Leptopterygius" (= Temnodontosaurus) platyodon. |
 Restoration | |
Sauropterygia
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A marine plesiosaur, member of the family Plesiosauria inside Sauropterygia. Non assigned to a concrete genus, probably are related to the Plesiosaurian genera from the Posidonia Shale |
||
|
|
|
A marine plesiosaur, member of the family Plesiosauria inside Sauropterygia. Non assigned to a concrete genus, probably are related to the Plesiosaurian genera from the Posidonia Shale |
||
|
|
|
A marine plesiosaur, member of the family Plesiosauridae inside Plesiosauroidea. Non assigned to a concrete genus, probably are related to the Plesiosaurian genus Microcleidus or to Seeleyosaurus, both know from Toarcian strata. |
||
|
|
Three articulated dorsal vertebrae |
A marine plesiosaur, member of the family Microcleididae inside Plesiosauroidea. Non assigned to a concrete genus, probably are related to the Plesiosaurian genus Microcleidus |
| |
|
|
|
A marine pliosauroid, member of the family Rhomaleosauridae inside Pliosauroidea. The genus is known from the Toarcian of southern Germany. |
| |


Crocodyliformes
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cervical vertebra |
A freshwater Crocodrilian, member of the family Goniopholididae inside Neosuchia. Non assigned to a concrete genus, originally labeled as Mesoeucrocodylia indet. can be the earliest representative of the group on Europe. Is considered to come from an osteologically immature individual. |
||
|
|
Partial rostrum with teeth |
A marine Crocodrylomorph, member of the family Thalattosuchia inside Neosuchia. Probably related to Pelagosaurus. The tooth morphology and the internal rostral anatomy indicate that it can be positively referred to a longistrine thalattosuchian crocodyliform. The size of PLM uncatalogued, as compared to the corresponding portions of the rostra of other Toarcian thalattosuchians, is indicative of a juvenile individual. |
||
|
|
Incomplete skull and associated osteoderm |
A marine Crocodrylomorph, member of the family Machimosauridae inside Teleosauroidea. A marine crocodylomorph with a diet probably based on fish.[136] It was identified as Steneosaurus. Although highly incomplete, the preserved maximum antorbital width of about 50 mm demonstrates that GG 422/6 can be clearly referred to an immature individual. |
| |

Theropoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Orionides? Indeterminate |
|
Dorsal Vertebrae |
A Saurischian, member of the family Orionides inside Tetanurae. The affinities of the Specimen aren't clear due to its fragmentary nature. Has been classified as Saurischia indeterminate, although shows clearly characters of the Orionides group (concave articular surfaces and a dished lateral pleurocoel, remnants of the neural arch and postzygapophyses).[137] The vertebrae centrum measures 80 mm, implying a medium-sized theropod (~5 m long).[137] Can be related with Yunyangosaurus. |
 Marshosaurus, example of basal Orionides | |
Sauropoda
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Gravisauria Indeterminate[140] |
|
|
A Saurischian, member of the family Gravisauria inside Sauropoda. The specimen is believed to be a juvenile, based on the ossification and unfused spine. Has affinities with the genus Tazoudasaurus and it is clearly distinctive form the also Toarcian Ohmdenosaurus, who is thought to be more basal.[140] The pelvic girdle elements can be clearly placed among the Sauropoda, on account of the presence of an elongated and strongly dorsally expanded iliac preacetabular process a possible relative.[140] The ischia GG411/3-4 resemble those of Tazoudasaurus in exhibiting a subtriangular iliac peduncle which create a short anteriorly directed expansion to reach the medial acetabular rim, and are slightly less developed than the Genus Barapasaurus.[140] |
 Barapasaurus can be a close relative of the Grimmen Sauropod | |
|
Gravisauria Indeterminate[140] |
|
|
A Saurischian, member of the family Gravisauria inside Sauropoda. The specimen is believed to be a juvenile (its comparatively small size is indicative of belonging to a not fully grown individual) and probably related to the Asian genus Zizhongosaurus, as shares characters with the neural spine V9067.1.[140] GG412 within Gravisauria is indicated by the presence of a well-developed spinodiapophyseal lamina.[140] |
||
Thyreophora
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Right side of the skull, the right lower jaw, caudal vertebrae, neural arches, a radius, a metatarsal, a claw, fragments of ribs, scutes and plates.[141] |
An Ornithischian, basal member of Thyreophora inside Genasauria. Its juvenile status makes controversial its phylogeny, being considered a relative of the also Early Jurassic Genus Scelidosaurus.[141] The compacted pile of disarticulated cranial and postcranial elements of the basal thyreophoran Emausaurus has been suspected to be a Speiballen (i.e., a compacted mass of indigestible stomach contents) regurgitated by a large marine reptile.[141] |
 Emausaurus attacked by a theropod | |
Plantae
Coals
.jpg.webp)
The Łęka Coal Basin has been known since 1800 for its abundant deposits; younger material has been suggested as redeposited from the Paleozoic, but the basin primarily yields a series of Pliensbachian-Toarcian coals.[143] In the Early Jurassic the Blanowice Formation was surrounded by land on the north, east and south, the source of sediment which was deposited as nearshore coal.[143][144] This coal is abundant in the upper part of the formation, dominated by alluvial and lacustrine sand and coal-bearing sediments.[145] Organic matter associated with the coal includes the oldest known biomolecules (in the Mrzygłód clay-pit), composed of labdanoic acid, ferruginol, sugiol and 7-oxototarol.[145] The extracted samples were recovered at the Wysoka Lelowska 47Ż and Jaworznik 124Ż boreholes; five core samples were taken from the Żarki 90Ż core, the latter from an approximately 1.5-metre (4 ft 11 in) coal seam.[145] The random reflectance (%Rr) of the coals is 0.47–0.56, indicating sub-bituminous coal.[145] The geochemical data suggest low-maturity sediments,[145] and hopane isomers are relatively high in all samples.[145] This is confirmed by unstable biomolecules in the coal samples, including labdanoic acid, ferruginol, sugiol, oxototarol, beta-sitosterol and cholesterol.[145]
The coals are generally dominated by vitrinite macerals, except for a high percentage of inertinite. This indicates wildfires or peat fires, confirmed by charcoal fragments.[145] Sesquiterpenes and diterpenes, common in conifers, angiosperms and bryophytes, were also recovered from the coal.[146] Vitrinite has a reflectance value of 0.49-0.56 %Ro. The cupressaceae and podocarpaceae families are considered the main peat-forming plant species, due to the presence of phenolic abietanes and dehydroabietic acids.[145] Lignite indicates significant benzohopane derivatives in the surrounding sandstones, probable differences in biodegradation, and a typically-low coalification range.[147] Later, larger studies note the influence of fires on the region.[148]
The Kaszewy coals, found in an approximately 150-metre (490 ft) section of terrestrial and marine siliciclastic sediments in the Kaszewy-1 and Niekłan PIG-1 boreholes, are the Ciechocinek Formation's major coals.[148] This section was in a nearshore-deltaic setting, with increased terrestrial and marine organic matter reflecting increased weathering and transport of terrestrial matter.[148] Abundant fossil charcoal and polycyclic aromatic hydrocarbons have been found. The number of coarse fossil charcoal particles (larger than 125 micrometers) in the Pliensbachian-Toarcian sections of the Kaszewy-1 core is very low (0-15 particles/10 g sediment), and fine charcoal particles (<125 μm) are more abundant (~12,000–256,000 particles/10 g sediment); there are also more non-charcoal particles.[148] There are more fine charcoal particles at the beginning of the Toarcian, reflecting environmental changes. In the polycyclic aromatic hydrocarbons, the pyrolytics (benz-anthracene, benzo(k)fluoranthene, fluoranthene, indeno[1,2,3-cd]pyrene, phenanthrene and pyrene) were detected in a wide variety of samples; phenanthrene is the most abundant component and coronene the least, suggesting the burning of organic matter.[148] Petrogenics are more abundant on the coal samples than pyrolytics, suggesting low wildfire activity.[148] Although the Kaszewy-1 borehole did not indicate increased wildfire activity, the fine fossil-charcoal abundance and pyrolytic concentration indicate regional wildfires.[148] Pyrolytics indicating the increased wildfire activity match the beginning of the Toarcian anoxic event, with intervals of fewer wildfires.[148] Wildfire changes match the Lower Toarcian negative carbon-isotopes emissions measured on the, which probably promoted a rise in atmospheric oxygen. Some questions remain; the climate was warmer and wetter (which can suppress wildfire activity), and wildfires persisted in the Kazewy-1 borehole wildfire activity was successfully sustained.[148] Wildfires may have subsided due to a lack of suitable fuel.[148]
Chlorophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Annulina[1] |
|
|
|
A marine Green Algae, member of the family Cladophoraceae inside Ulvophyceae. A genus found related with a high diversity of marine Foranimifers, and mostly linked with the pomeranian deposits. It was the only identified Algae, and its fossils were confused with calcareous Foranimifer cunchs. |
 Extant Chaetomorpha, the typical example of the Cladophoraceae. Annulina probably come from a similar Green Algae, tougth it may have also come from microscopic specimens |
Megaspores
Polish Lower Toarcian palynology is assigned to the Paxillitriletes phyllicus (Ph) level (Isoetales), due to the abundance of this genus.[149] The lower part of the Toarcian level has numerous occurrences of this species, sometimes before the genera Erlansonisporites sparassis (Selaginella-like) and Minerisporites volucris (Isoetaceae) in the Gorzów Wlkp. IG 1 borehole.[149] The upper part contains less of the genus.[149] The most common species found on Poland in this era include Erlansonisporites sparassis, E. excavatus, Minerisporites volucris and Biharisporites scaber (Lycopodiopsida); Aneuletes potera and Trileites murrayi (both Selaginella) are found on the upper levels.[149] The Toarcian disturbance of the carbon cycle recorded on the Ciechocinek Formation, coincides roughly with the appearance of Paxillitriletes phyllicus. The type of dominant palynomorphs recovered changed from pollen grains during the Upper Pliensbachian to megaspores, indicating a climatic change from moderate and relatively dry to warm and humid in the early Toarcian.[149] This shift in local climate correlates with a global maritime transgression in which volcanism in the Karoo-Ferrar large igneous provinces raised the global temperature and disrupted the carbon cycle, creating a major greenhouse effect.[149] The prevalence of megaspore Paxillitriletes phyllicus correlate with warmth and humidity; the flora, dominated by the family Isoetaceae, requires standing water to reproduce.[149] The megaespore Paxillitriletes phyllicus then drops significantly, indicating a return to a more moderate climate during sedimentation of the younger Borucice Formation.[149]
Bryophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Sphagnopsida inside Sphagnales. Spores nearly identical to that one found associated with the modern moss genus Sphagnum. Moss related to high humid environments. |
 Extant Sphagnum, the typical example of the Sphagnales. Rogalskaiporites probably come from a similar Plant | |
|
Cingulatisporites[151] |
|
|
|
Affinities with Anthocerotaceae ( specially Phaeoceros, Megaceros) inside Bryophyta. Moss spores from humid settings |
_1077.JPG.webp) Extant Phaeoceros, whose spores are similar to the extinct Cingulatisporites |
|
Cirratriradites[151] |
|
|
|
Affinities with Anthocerotaceae inside Bryophyta. Moss spores from humid settings |
 Extant Anthoceros, the typical example of the Anthocerotaceae. Cirratriradites probably come from a similar Plant |
Lycophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Lycophyta inside Tracheophyta. A relative abundant genus, specially on deltaic deposits. Low herbaceous Lycophyte flora. |
_002.jpg.webp) Extant Lycopodium, typical example of Lycophyta. Bacutriletes probably come from a similar or a related Plant | |
|
Acanthotriletes[154] |
|
|
|
Affinities with Lycophyta inside Tracheophyta. These spores, rather abundant are correlated with humid settings, and come from mostly herbaceous-arbustive plants. |
 Extant Lycopodium, typical example of Lycophyta. Acanthotriletes probably come from a similar or a related Plant |
|
|
|
Affinities with Lycophyta inside Tracheophyta. Spores from low herbaceous flora, linked mostly to humid environments with abundant freshwater. |
 Extant Lycopodium, typical example of Lycophyta. Hughesisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycophyta inside Tracheophyta. Low herbaceous flora from humid envrironments |
 Extant Lycopodium, typical example of Lycophyta. Eehitriletes probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycophyta inside Tracheophyta. Rather rare than other similar genera, found mostly on deltaic facies. |
 Extant Austrolycopodium, typical example of Lycophyta. Thuringiasporites probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycopodiaceae inside Lycopodiopsida. Represents herbaceous Lycophytes of small to medium size (10–40 cm), that are found mostly on deltaic deposits. |
_(25044755342).jpg.webp) Extant Lycopodium, typical example of Lycopodiaceae. Trachytriletes probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycopodiaceae inside Lycopodiopsida. |
.jpg.webp) Extant Diphasiastrum, typical example of Lycopodiaceae. Raistrickia probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycopodiaceae inside Lycopodiopsida. Resemble spores of the modern genus Lycopodium. If it belongs to a similar genus, represent low herbaceous flora spores. |
 Extant Lycopodium, typical example of Lycopodiaceae. Lycopodiumsporites probably come from a similar or a related Plant | |
|
|
|
Affinities with Lycopodiaceae inside Lycopodiopsida. Resembles the modern genera Lycopodiella and Huperzia, characteristic small to medium (20–90 cm) herbaceous Lycopites, found on environments with abundant water. |
 Extant Huperzia, typical example of Lycopodiaceae. Reticulatisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Lycopodiopsida inside Lycophyta. Lycophyte spores of rather uncertain affinities, are more common on fluvial deposits. |
 Extant Huperzia, typical example of Lycopodiaceae. Biharisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Isoetales inside Lycophyta. Spores related to modern Isoetes, representing Small plants related to water bodies.[160] It comprises the main Megaspore zonation of the Toarcian of Poland, being the most abundant spore found on the Ciechocinek Formation, and the genus that marks the start of the strata. The Abundance of this genus along with Minerisporites institus points to very humid conditions during almost the entire T-OAE, upwards from CIE step 2 of the carbon isotope curves. |
| |
|
|
|
Affinities with Isoetaceae inside Lycopsida. Related to plant similar to Isoetes lacustris, present on flooded basins, and other ecosystems with relative abundant water supply.[161] Minerisporites institus is the second most abundant genus of spore found on the Ciechocinek Formation. |
| |
|
|
|
Affinities with Isoetaceae inside Lycopsida. |
| |
|
|
|
Affinities with the genus Selaginella inside Selaginellaceae. Herbaceous Moss.[163] Relatively abundant Pollen Genera |
 Extant Selaginella. Erlansonisporites spores are similar to the ones on this genus | |
|
|
|
Affinities with Selaginellaceae inside Lycopsida. A genus more common on younger strata, relatively more abundant than similar genera, found mostly on deltaic facies. |
 Extant Selaginella, typical example of Selaginellaceae. Aneuletes probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Less abundant than othe Lycophite Spores, its abundance increases near Deltaic deposits. |
 Extant Selaginella, typical example of Selaginellaceae. Horstisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. |
 Extant Selaginella, typical example of Selaginellaceae. Hymenozonotriletes probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. The most diverse palynological residue on the formation, the 3rd most abundant genera of Spore present on the Formation and one of the most abundant on the south of Fennoscandia |
 Extant Selaginella, typical example of Selaginellaceae. Trileites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Among the most abundant genera of Pollen present on the Formation and the south of Fennoscandia |
 Extant Selaginella, typical example of Selaginellaceae. Triletes probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Among the most abundant genera of Pollen present on the Formation and the south of Fennoscandia |
.jpg.webp) Extant Selaginella, typical example of Selaginellaceae. Lagenicula probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. This herbaceous Lycophyte flora appears more on the southern boreholes, linked to large wood debris. Resemble extant Selaginella erthropus |
 Extant Selaginella, typical example of Selaginellaceae. Valvisisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Herbaceous flora spores linked to deltaic facies. Resemble extant Selaginella erthropus |
 Extant Selaginella, typical example of Selaginellaceae. Maexisporites probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Resemble extant Selaginella erthropus |
 Extant Selaginella, typical example of Selaginellaceae. Striatriletes probably come from a similar or a related Plant | |
|
|
|
Affinities with the Selaginellaceae inside Lycopsida. Less abundant than other similar spores, found on a few samples on all the boreholes. Altrougth is quite diverse. |
.JPG.webp) Extant Selaginella, typical example of Selaginellaceae. Foveosporites probably come from a similar or a related Plant | |
|
|
|
Affinities with Pleuromeiales inside Lycophyta. The Plueromeiales were tall Lycophites (2 to 6 m) common on the Trassic. Probably come from a relict genus. |
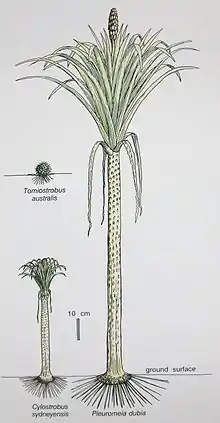 Reconstruction of the extinct genus Pleuromeia, typical example of Pleuromeiales. Nathorstisporites probably come from a similar or a related Plant | |
|
Margaritatisporites[151] |
|
|
|
Affinities with Pleuromeiales inside Lycophyta. |
|
|
Bacutriletes[151] |
|
|
|
Affinities with Pleuromeiales inside Lycophyta. |
|
|
Crassispora[151] |
|
|
|
Affinities with Sigillariaceae inside Lycophyta. Arboreal Lycopites |
|


.jpg.webp)
Equisetidae
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Asterocalamotriletes[151] |
|
|
|
Affinities with Asterocalamitopsida inside Equisetopsida. Is related with the Palaeozoic horsetails Asterocalamites and Stigmatocanna. Tall Horsetails of up to 4 m. |
|
|
|
|
Affinities with Equisetopsida inside Equisetopsida. Typical Horsetail Spores. |
 Extant Equisetum, the typical example of the Equisetopsida. The "Equisetum Spores" are pretty similar to the extant ones found on this genus | |
|
Florinisporites[151] |
|
|
|
Affinities with Equisetopsida inside Equisetopsida. Found only on Poland. |
.JPG.webp) Extant Equisetum cone, the typical example of the Equisetopsida. Florinisporites spores are pretty similar to the extant ones |
|
Irrimales[151] |
|
|
|
Affinities with Equisetopsida inside Equisetopsida. Found only on Poland. |
 Extant Equisetum cone, the typical example of the Equisetopsida. Irrimales spores are pretty similar to the extant ones |
Gymnospermopsida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Gymnospermopsida inside Tracheophyta. Pollen that resemble those of paleozoic genera such as Gangamopteris, with arboreal built. |
||
|
|
|
Affinities with Gymnospermopsida inside Tracheophyta. Pollen that resemble those of paleozoic genera such as Gangamopteris, with arboreal built. |
||
|
|
|
Affinities with Gymnospermopsida inside Tracheophyta. Pollen that resemble those of paleozoic genera such as Gangamopteris, with arboreal built. |
||
Filicopsida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the Filicopsida inside Monilophyta. Fern spores of uncertain placement |
||
|
|
|
Affinities with the Filicopsida inside Monilophyta. Fern spores of uncertain placement |
||
|
|
|
Affinities with the Polypodiidae inside Filicopsida. Fern spores of uncertain placement. |
||
|
|
|
Affinities with the Polypodiidae inside Filicopsida. Fern spores of uncertain placement |
||
|
|
|
Affinities with the Schizaeaceae inside Schizaeales. Represents large spores from herbaceous ferns, that are more abundant on the south of the Toarcian polish basin than on the north. |
 Extant Schizaea, typical example of the Schizaeaceae. Klukisporites spores are pretty similar to the extant ones of this genus | |
|
|
|
Affinities with the Cynepteridaceae inside Schizaeaceae. Related to Ferns similar to Cynepteris, from the late triassic of North America. |
||
|
|
|
Affinities with Lygodiaceae inside Schizaeales. Spores nearly identical to that one found associated with the modern moss genus Lygodium. Arboreal moss related to high humid environments, being major fuel for peat fires. |
 Extant Lygodium, typical example of the Lygodiaceae. Reticulatisporites spores are pretty similar to the extant ones of this genus | |
|
|
|
Affinities with the Aspleniaceae inside Filicopsida. |
 Extant Asplenium, typical example of the Aspleniaceae | |
|
Laevigatosporites[151] |
|
|
|
Affinities with the Blechnaceae inside Filicopsida. |
 Extant Sadleria, typical example of the Blechnaceae. Laevigatosporites spores are pretty similar to the extant ones of this genus |
|
Dennstaedtiites[151] |
|
|
Spores |
Affinities with Dennstaedtiaceae inside Pteridophyta. Floor Herbaceous ferns, probably from covered forest areas. |
.jpg.webp) Extant Pteridium aquilinum, typical example of the Dennstaedtiaceae. Dennstaedtiites spores are pretty similar to the extant ones of this genus |
|
|
Spores |
Affinities with Dennstaedtiaceae inside Pteridophyta. |
 Extant Onychium. Some local spores are similar to the ones found on this genus | |
|
Baculatisporites[151] |
|
|
Spores |
Affinities with Osmundaceae (specially Osmunda and Leptopteris) or Hymenophyllaceae (specially Hymenophyllum cruentum) inside Pteridophyta. |
 Extant Hymenophyllum, typical example of the Hymenophyllaceae. Baculatisporites may have come from a similar genus. Alternatively it can be spores from an Osmundaceae fern |
|
Todites[151] |
|
|
Spores |
Affinities with Osmundaceae (specially Leptopteris) inside Pteridophyta. The most abundant Fern Spore found on all the Formation |
 |
|
|
Spores |
Affinities with Osmundaceae inside Pteridophyta. Spores nearly identical to that one found associated with the modern fern genus Osmunda. Members of the genus Osmunda have been found on coeval age strata on Sweden. |
 Extant Osmunda, typical example of the Osmundaceae. Osmundacidites may have come from a similar genus | |
|
|
|
Affinities with the Marsileaceae inside Salviniales. Represents spore from fully acuatic ferns, found associated with fluvial or deltaic deposits, where probably formed large underwater colonies. |
 Extant Marsilea, typical example of the Marsileaceae. Crybelosporites may have come from a similar genus | |
|
|
|
Affinities with the Gleicheniaceae inside Polypodiidae. Resemble the modern Gleichenia Spores, and proably represent a similar genus or a member of it. Fern related to large colonies, found mostly on humid environments. |
 Extant Unidentified Gleicheniaceae. Gleicheniidites may have come from a similar genus | |
|
|
|
Affinities with the Marattiaceae inside Polypodiidae. Resemble the modern Marattia spores, probably belonging to a similar genus, related with large sized herbaceous ferns of humid environments. |
 Extant Marattia, typical example of the Marattiaceae. Marattisporites may have come from a similar genus | |
|
|
|
Affinities with the Marattiaceae inside Polypodiidae. |
 Extant Marattia, typical example of the Marattiaceae. Camptotriletes may have come from a similar genus | |
|
|
|
Affinities with the Matoniaceae inside Polypodiidae. The so-called comb-ferns, found forming large colonies on humid settings. |
 Extant Matonia, typical example of the Matoniaceae. Matonisporites may have come from a similar genus | |
|
|
Spores |
Affinities with Cyatheaceae inside Cyatheales. Cyathidites minor almost certainly belong to well known Mesozoic species Coniopteris hymenophylloides and to other fossil cyatheaceous or dicksoniaceous ferns such as Eboracia lobifolia and Dicksonia mariopteri. |
_(14820801765).jpg.webp) | |
Cycadeoidophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Bennettitinae[151] |
|
|
|
Affinities with Bennettitinae the inside Bennettitales. Very abundant Pollen. |
|
|
|
|
Affinities with the Cycadeoidaceae inside Bennettitales. Pollen identified originally coming from Pinaceae Conifers, was later proved to come from Bennetites. |
||
|
|
|
Affinities with the Cycadeoidaceae inside Bennettitales. |
||
|
|
|
Affinities with Williamsoniaceae the inside Bennettitales. Very abundant Pollen. |
 Reconstruction of Williamsonia, typical example of the Williamsoniaceae. Bennetistemon Pollen is considered to come from a similar genus | |
|
|
Pollen |
Named originally Pollenites apertus, they resemble pollen grains of the genus Cycas. Later works refer them to modern pollen grains that proper of wrote that resemble ?Cycadopsida (?Cycadales). Alternatively, can be Pollen from members of Ginkgopsida (?Gnetales).[164] |
 Extant Encephalartos, typical example of the Cycas. Chasmatosporites Pollen is pretty similar to the extant ones of this genus. Alternatively, it maybe come from Ginkos or Gnetales | |
Gnetophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Gnetopsida and probably Gnetophyta. Has Been considered Pollen of Chloranthaceae. However, it is to old for belonging to advanced Angiosperms. It probably comes from cones related to the Genera Piroconites kuesperti from the Lowermost Jurassic of Germany, resembling pollen of extant Ephedra and Welwitschia. |
 Closer Look of Ephedra cones, a common Gnetal. Clavatipollenites maybe come from a related plant | |
|
Ephedripites[151] |
|
|
|
Affinities with the Chlamydospermae inside Gnetales. Identified originally as Pollen coming from the extant Ginkgo biloba, was later found to be misidentified. It was latter foun that Aphlebia lautneri from the Hettangian of Franken (Germany) has microsporophylls covered on one surface with synangia consisting of three adnate pollen sacs, yielding Ephedripites pollen.[165] |
|
|
|
|
Type Pollen of the Erdtmanithecales, that can be related with the Gnetales. Thick tectum, infratectum of small granules, indistinct or absent foot layer. Originally was thought to come from Angiosperms, latter reports suggest it come from arbustive Bennetites.[166] It was recently found to come from Eucommiitheca, member of the enigmatic Erdtmanithecales, reinterpreted as an unusual gymnosperm grain with a single distal colpus flanked by two subsidiary lateral colps.[167] Is very similar to the Pollen of the extant Ephedra and Welwitschia (mainly on the granular structure of the exine).[168] |
||

Coniferophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Voltziales inside Coniferae. Primitive Conifer and possible relict taxon |
||
|
|
|
Affinities with Voltziales inside Coniferae. Primitive Conifer and possible relict taxon |
||
|
|
|
Affinities with Paleoconiferae inside Coniferae. Primitive Conifer and possible relict taxon |
||
|
Protoabies[151] |
|
|
|
Affinities with Protoconiferae inside Coniferae. |
|
|
|
|
Affinities with Abietoideae inside Coniferae. |
.jpg.webp) Extant Abies Cone, example of the Abietoideae. Some Pollen recovered on the Breholes of the formation is similar to the one found on this genus | |
|
|
|
Affinities with Abietoideae inside Coniferae. Pollen From arbustive to arboreal plants, resembling the pollen of the modern genus Tsuga. The differences observed between Cerebropollenites and Tsuga are no greater than the differences observed between the pollen of the two Sections of Tsuga, Hesperopeuce and Micropeuce. |
 Extant Tsuga Cone, example of the Abietoideae. Cerebropollenites is similar to the pollen found on this genus | |
|
|
Pollen |
Affinities with Abietoideae inside Coniferae. Pollen From arbustive to arboreal plants, resembling the pollen of the modern genus Tsuga |
 Extant Tsuga Cone, example of the Abietoideae. Tsugaepollenites is similar to the pollen found on this genus, hence the name | |
|
|
|
Affinities with the Pinidae inside Coniferae. |
 Extant Pinus cembra Cone, example of the Pinidae. Inaperturopollenites is similar to the pollen found on this genus | |
|
|
|
Affinities with the Pinidae inside Coniferae. |
| |
|
Rotundipollenites[154] |
|
|
|
Affinities with Pinaceae inside Coniferae. |
|
|
|
|
Affinities with Pinaceae inside Coniferae. Resemble modern Pinus Pollen, probably belonging to a similar Genus. |
| |
|
|
|
Affinities with Pinaceae inside Coniferae. Pollen From arbustive to arboreal plants, resembling the modern genus Picea |
| |
|
|
Pollen |
Affinities with Sciadopityaceae inside Coniferae. This pollen resembles the present on the modern Cupressaceae Sciadopitys |
 Extant Sciadopitys Cone, example of the Sciadopityaceae. Sciadopityspollenites is similar to the pollen found on this genus | |
|
|
|
Affinities with Podocarpaceae inside Pinopsida. Pollen From arbustive to arboreal plants |
 Extant Podocarpus Cone, example of the Podocarpaceae. Pseudopodocarpus is similar to the pollen found on this genus | |
|
|
Pollen |
Affinities with Podocarpaceae inside Pinopsida. Pollen From arbustive to arboreal plants. Pollen that resemble the Pollen of the extant genus Prumnopitys taxifolia |
 Extant Prumnopitys Cone, example of the Podocarpaceae. Pristinuspollenites is similar to the pollen found on this genus | |
|
|
|
Affinities with the Cheirolepidiaceae inside Coniferae. Is very abundant on hot and dry settings, found specially on the Brody-Lubienia Borehole. |
||
|
|
|
Affinities with the Cupressaceae inside Coniferae. The Pollen from this genus is similar to the present on the modern Fitzroya and Calocedrus. |
 Extant Calocedrus Cone, example of the Cupressaceae. Cupressacites is similar to the pollen found on this genus | |
|
|
|
Affinities with the Araucariaceae inside Coniferae. Resemble the pollen from the modern genus Agathis. |
 Extant Agathis Cone, example of the Araucariaceae. Applanopsipollenites is similar to the pollen found on this genus | |
|
|
|
Affinities with the Araucariaceae inside Coniferae. Resemble the pollen from the modern genus Agathis. |
 Extant Agathis. Some Pollen recovered on the Boreholes is similar to the one found on this genus | |
|
|
Pollen |
Affinities with Araucariaceae inside Pinopsida. Pollen From arbustive to arboreal plants |
 Extant Araucaria Cone, example of the Araucariaceae. Araucariacites is similar to the pollen found on this genus | |
.03.jpg.webp)



Fossil wood
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with the family Ginkgoaceae inside Ginkgoopsida. It is scarce, and shows characters similar to the Baieroxylon specimens found on coevel strata from France and Germany. Represents wood from Arboreal Ginkgo-like plants. The upper part of the Carbon deck at Kierszula is characterized by large lignites, where it is thicker (0.9–4.6 cm) streaks or lenses often show clear structural features of wood on the surfaces perpendicular to the stratification. |
 | |
|
|
|
Affinities with the family Cheirolepidiaceae inside Coniferales. Generally, the most abundant Genus of Wood recovered locally on the Formation. Most of the wood studied from the Lower Toarcian of Poland, recovered on the Gorzów Wielkopolski Borehole come from this genus, that has revelated the effect of the Toarcian Oceanic Anoxic event over the local wood, where the more humid local climate led this genus of wood to grow on restricted nearshore marine through to barrier-lagoon environments, hence the abundance of trunks.[33] |
||
|
|
|
Affinities with the family Cheirolepidiaceae inside Coniferales. It is locally the second most abundant genus of Wood, widespread across Europe on the Lower Jurassic.[33] Some Cuticles resemble the structure seen on the invalid species Brachyoxylon rotnaense (Syn. of Simplicioxylon), present on the Lower Liassic of Denmark. This genus is known from abundant Xylogic Material from Coeval deposits of Hungary. |
||
|
|
|
Affinities with the family Araucariaceae inside Coniferales. Represents the major type of Wood found on the Kierszula coals (up to 55%). This genus is also widely populated by different sorts of parasite fungus, specially on Wood cuticles found after the Toacian Anoxic event boundary.[11] |
 | |
|
|
|
Affinities with Coniferales, that can be a member of the extinct family Miroviaceae. Was found to be close to the Podocarpaceae, Cupressaceae and in a lesser extent to the Cheirolepidiaceae. Xenoxylon co-occurred with Sciadopitys-like foliages in Siberia (Tritaenia and Sciadopityoides, typical Miroviaceae), and the type genus Mirovia szaferi is known from the Bathonian of Poland.[170] This genus is marker for a cold and/or humid climate, being probably the main reason is mainly absence on mostly of the Wood samples, being rather rare, as the Ciechocinek Formation deposited on Humid and Hot climate.[169] |
||
Megaflora
The Lublin upland fluvial sandstones contain diverse fossil flora, associated genera and species with Lower Jurassic sediments. Carboniferous flora, similar to that of Jurassic formations, appeared in boreholes near the planned Bogdanka Coal Mine.[171] The age of the plant material was not determined until 2020, when it was identified as Lower Toarcian (with some Late Pliensbachian strata).[172] The Brody-Lubienia borehole is abundant in terrestrial palynomorphs and aquatic biomass.[172] The sediments from Brody-Lubienia have a more-terrestrial character, indicated by the frequent occurrence of plant roots and paleosol horizons;[172] moisture was probably fresh (not saline) water.[172] The environment was probably dry, developing flora near freshwater inflow from the east.[172]
The Lublin lias is dominated by cycads and Bennetites Ginkgoales. Ferns occur sporadically in the bottom of the Toarcian, where deposits are filled with coal, mudstone, sandstone and clay siderite (reworked from the Carboniferous) and pebbles from Devonian limestones.[173] Similar boreholes and nearby deposits indicate the end of a river which transported Devonian-Carboniferous deposits from the northeast to the aquatic inland environment.[173] Vegetation grew primarily outside the sedimentation area, on shores and in shallow water.[174]
Equisetopsida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Stems |
Affinities with Equisetaceae inside Equisetales. Equisetalean Stems, related that are also found on the Hettangian strata along Skane, Sweden. The washed Equisetalean stems are the most abundant flora recovered on the German Realm of the Formation. |
||
|
|
|
Affinities with Equisetales inside Equisetopsida. This genus is rather scarce compared with other coeval Equisetalean fossils |
 | |
|
|
|
Affinities with Calamitaceae inside Equisetopsida. A common horsetail on the Liassic of Europe. . |
 | |
|
|
Leaf Whorls |
Affinities with Equisetales inside Equisetopsida. Only a few fragmentary leaf whorl impressions of Equisetalean plants have been discovered, its description is nearly impossible as are not complete enough for a concrete diagnosis. |
||
Selaginellaceae
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
Selaginellites[179] |
|
|
Leafy Branch |
Affinities with Selaginellaceae inside Selaginellales. A Fragmentary Specimen was recovered, but its status is rather controversial. It comes from a series of laminated Sandstone/Mudstone levels on the profil of Przysucha with numerous clusters of muscovite and cletrite and other charred plants, probably derived from a freshwater setting. |
|
Pteridospermatophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Pinnae |
Affinities with the Peltaspermales inside Pteridospermatophyta. Bipinnate leaves, rachis longitudinally striated, with a long petiole and secondary rachises inserted oppositely to suboppositely on the upper side of the primary rachis. This Leaves belong to large Aboreal Ferns related with dry environments. |
||
|
|
Pinnae |
Affinities with the Umkomasiaceae inside Pteridospermatophyta. Is very similar to Rhaphidopteris, characterized by usually bi-tripinnate, coriaceous leaves with narrow segments. It belongs to Large (up to 25 m tall) arboreal Fern-like plants. |
||
|
|
Ovulate Structure |
Affinities with the Caytoniaceae inside Caytoniales. Represents the Ovulate organs or large Tree ferns, and is related with the middle Jurassic flora of United Kingdom. |
 | |
|
|
Pinnae |
Affinities with the Caytoniaceae inside Caytoniales. Sagenopteris is most likely the Pinnae of the Tree Fern that also contain Caytonia. |
 | |
Bennettitopsida
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. Anomozamites is characterised by slender, (almost) completely and regularly segmented leaves whose leaflets are generally as long as broad or, at maximum, two times as long as broad. This genus is related with more arboreal Bennetitalean flora. Shows coriaceous leaves and is a genus linked more with dry climates. |
 | |
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. A more arbustive type of Bennetite, abundant on the Pliensbachian-Toarcian strata along Eurasia, related to rather dry climates. |
.jpg.webp) | |
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. This Bennetite has a leaf similar to Nilssonia, large, lanceolate in outline with coriaceous structure, like the modern angiosperm Philodendron. Is related with rather Dry-hot environments, with less Fern flora. |
||
|
|
Pinnae |
Affinities with the Cycadeoidaceae inside Bennettitales. The most abundant plant macrofossil locally, and the most diverse found on the Lublin coals. It represents a Cycad Like plant with Coriaceous leaves. Is related by some botanists with the Bennetite branch that led to Angiosperms. |
 | |
|
|
Bennetite "Flower" |
Affinities with the Cycadeoidaceae inside Bennettitales. The Bennetite flowers are the main organ that links this relatives with the Cycas with modern Angiosperms. Probably the Pollen assigned to Angiosperms come from plants with this "Flowers". |
||
|
|
Pinnae |
Affinities with the Williamsoniaceae inside Bennettitopsida. Leaves from Arboreal Bennetites, similar to the modern Cyca Encephalartos woodii, with robust Trunks, built for Dry and hot climates. |
 | |
Ginkgoidae
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Leaves |
Affinities with the Ginkgoaceae inside Ginkgoidae. Linked to the Hettangian-Sinemurian flora from Greenland and Skane, but also with coeval flora from the Sorthat Formation. Is the main Tree flora recovered locally. |
 | |
|
|
Cuticles |
Affinities with the Ginkgoaceae inside Ginkgoidae. Associated with relatively warmer and humid conditions. |
 | |
|
|
Cuticles |
Affinities with the Ginkgoaceae inside Ginkgoidae. Associated with relatively warmer and humid conditions. |
||
Coniferophyta
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cone Scales |
Affinities with the Cycadocarpidiaceae inside Pinales. The only non Equisetalean plant megafossil from the German realm, is represented by a few cone scales and related with the Lower Jurassic Flora of Scania. Ulmatolepis bears a striking resemblance to Cycadocarpidium, and correlates this flora with the Shasonggang of Jilin in the Northern Palaeofloristic Province of China. |
||
|
|
Ovuliferous cones |
Affinities with the Cheirolepidiaceae inside Pinales. The type Leave from the Cheirolepidaceae family (as Cheirolepis, the genus that give name to the family, is a junior synonym), appears to be linked with Hot climates, able to survive in dry, extreme conditions, and been fire tolerant. |
||
Footnotes
- Kopik, J. (1960). Mikropaleontologiczna charakterystyka liasu dolnego doggeru Polski. Geological Quarterly, 4(4), 921–935.
- Jurkiewiczowa, I. (1967). Lias zachodniego obrzeżenia Gór Świętokrzyskich i jego paralelizacja z liasem Wyżyny Krakowsko-Częstochowskiej. Biul. Inst. Geol, 200, 5-132.
- Osika, R. (1958). Profil górnego liasu i doggeru okolic Złotowa. Geological Quarterly, 2(4), 765–784.
- Kopik J. and Marcinkiewicz T. 1997. Jura dolna: Biostratygrafia. In: S. Marek and M. Pajchlowa (Eds), The epicontinental Permian and Mesozoic in Poland (in Polish with English summary). Prace Paƒstwowego Instytutu Geologicznego, 153: 196-205.
- Kopik, J. (1998). Lower and Middle Jurassic of the north-eastern margin of the Upper Silesian Coal Basin. Biuletyn Państwowego Instytutu Geologicznego, 378, 67–129.
- Nagy, J., Hess, S., & Alve, E. (2010). Environmental significance of foraminiferal assemblages dominated by small-sized Ammodiscus and Trochammina in Triassic and Jurassic delta-influenced deposits. Earth-Science Reviews, 99(1-2), 31–49.
- Jurkiewicz, H. (1965). Profil wiercenia w Bożej Woli. Przegląd Geologiczny, 13(9), 378.
- ZESSIN, W. (2010). Der Dobbertiner Jura (Lias ε, Mecklenburg) und seine Bedeutung für die Paläoentomologie. Virgo, Mitteilungsblatt des Entomologischen Vereins Mecklenburg, 13(1), 4–9.
- Franke, A. (1936). Die foraminiferen des deutschen Lias (No. 169). Im Vertrieb bei der Preussischen Geologischen Landesanstalt.
- Barski, M., & Leonowicz, P. (2002). Dinoflagellates of Lower Jurassic outcrops at Kozłowice and Boroszów (southern Poland). Przegląd Geologiczny, 50(5), 411–414.
- Pieńkowski, G., Hodbod, M., & Ullmann, C. V. (2016). Fungal decomposition of terrestrial organic matter accelerated Early Jurassic climate warming. Scientific reports, 6, 31930.
- Rogalska,M. (1954) Spore and pollen analysis of the Liassic Coal of Blanowice in Upper Silesia [ Analyza sporowo-pylkowa Liasowego Wegla Blano-Wickiego z Gornego Slaska ] Biuletyn - Instytutu Geologiczny Vol. 89 pp. 1–46
- F. E. Geinitz. 1880. Der Jura von Dobbertin in Mecklenburg und seine Versteinerungen. Zeitschrift der Deutschen Geologischen Gesellschaft 32:510-535
- SUHR, P. (1988). Lebensspuren aus dem Lias von Dobbertin. Fundgrube, 24(1), 22–26.
- Osgood, R. G. (1975). The history of invertebrate ichnology. The study of trace fossils (pp. 3–12). Springer, Berlin, Heidelberg.
- Hertweck, G., Wehrmann, A., & Liebezeit, G. (2007). Bioturbation structures of polychaetes in modern shallow marine environments and their analogs to Chondrites group traces. Paleogeography, Palaeoclimatology, Palaeoecology, 245(3-4), 382–389.
- Leonowicz, P. (2008). Trace fossils from the Lower Jurassic Ciechocinek Formation, SW Poland. Volumina Jurassica, 6(6), 89–98.
- Keighley, D. G., & Pickerill, R. K. (1995). Commentary: the ichnotaxa Palaeophycus and Planolites: historical perspectives and recommendations.
- De Gibert, J. M., & Benner, J. S. (2002). The trace fossil" Gyrochorte": etiology and paleoecology. Revista Espanola de paleontologia, 17(1), 1–12.
- Leonowicz, P. M. (2016). Tubular tempestites from Jurassic mudstones of southern Poland. Geological Quarterly, 60(2), 385-394.
- Czapowski, G., Dadlez, R., Feldman-Olszewska, A., Gortyńska, S., Jaskowiak-Schoeneichowa, M., Kasiński, J. R., ... & Znosko, J. (2014). Szczegółowy profil litologiczno-stratygraficzny.
- Chakraborty, A., & Bhattacharya, H. N. (2013). Spreiten burrows: a model based study on Diplocraterion parallelum. Geospectrum, 296–299.
- Ekdale, A. A., & Lewis, D. W. (1991). Trace fossils and paleoenvironmental control of ichnofacies in a late Quaternary gravel and loess fan delta complex, New Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology, 81(3-4), 253–279. doi:10.1016/0031-0182(91)90150-p
- Marcinkiewicz,T. (1971) The stratigraphy of the Rhaetian and Liasin Poland based on megaspore investigations [ Stratygrafia Retyku i Liasu w Polsce na Podstawie badan megasporowych. ] Prace Instytut Geologiczny,(Warsaw) Vol. 65 P. 1–57
- Manum, S. B., Bose, M. N., & Sawyer, R. T. (1992). Seeds (Burejospermum Krassilov) and palynomorphs (Dictyothylakos Horst) with a netted wall structure reinterpreted: Clitellate cocoons. Courier Forschungsinstitut Senckenberg, 147, 399-404.
- Denckmann, A. (1887). Ueber die geognostischen Verhältnisse der Umgegend von Dörnten nördlich Goslar: mit besonderer Berücksichtigung der Fauna des oberen Lias (Vol. 8, No. 2). In Commission bei der S. Schropp'schen Hof-Landkartenhandlung (JH Neumann).
- Suhr, P. (1988). Lebensspuren aus dem Lias von Dobbertin. Fundgrube, Bd.1, S.22.
- Wittern, A. (2008). Fossilien als Glazialgeschiebe in Norddeutschland III: Pern, Trias, Jura. EGMA-News, Jg.2008, H.1, S.10–17.
- SAMSONOWICZ J. (1929) - Cechsztyn, trias i lias na północnym obrżeżeniu Łysogór. Spraw. Państ. Inst. Geol, 5, nr 1-2, p. ;1–282. Warszawa.
- KRAJEWSKI R. (1947) - Złoża żelaziaków ilastych we wschodniej części powiatu koneckiego. Biul. Państw. Inst. GeoL, 26. Warszawa.
- KSIĄZKIEWICZ M., SAMSONOWICZ · J. (1952) - Zarys geolcigii Polski. PWN, pp. 90–130. Warszawa
- Kopik, J. (1960). O kilku morskich małżach z serii gielniowskiej liasu Gór Świętokrzyskich [About several sea clams from the series of Gielniowska lias of the Świętokrzyskie Mountains]. Geological Quarterly , 4 (1), 95–104.
- Hesselbo, S. P., & Pieńkowski, G. (2011). Stepwise atmospheric carbon-isotope excursion during the Toarcian oceanic anoxic event (Early Jurassic, Polish Basin). Earth and Planetary Science Letters, 301(1-2), 365–372.
- Dadlez, R. (1958). Badania geologiczne na antyklinorium pomorskim w roku 1957. Geological Quarterly, 2(4), 740–764.
- SAMSONOWICZ J. (1934) - Objaśnienia arkusza Opatów. Ogólna mapa geologiczna Poląkiw skali 1: 100000. Państw. Inst. GeoL Warszawa
- Teichert, S., & Nützel, A. (2015). Early Jurassic anoxia triggered the evolution of the oldest holoplanktonic gastropod Coelodiscus minutus by means of heterochrony. Acta Palaeontologica Polonica, 60(2), 269–276.
- GRÜNDEL, J. (1999a): Gastropoden aus dem höheren Lias von Grimmen, Vorpommern (Deutschland). – Archiv für Geschiebekunde, 2: 629–672.
- Lehmann, U. (1967). Ammoniten mit Kieferapparat und Radula aus Lias-Geschieben. Paläontologische Zeitschrift, 41(1-2), 38–45.
- Howarth, M. K., & Rawson, P. F. (1965).The Liassic succession in a clay pit at Kirton in Lindsey, north Lincolnshire. Geological Magazine, 102(3), 261–266.
- van de Schootbrugge, B., McArthur, J. M., Bailey, T. R., Rosenthal, Y., Wright, J. D., & Miller, K. G. (2005). Toarcian oceanic anoxic event: An assessment of global causes using belemnite C isotope records. Paleoceanography, 20(3).
- Ansorge, J. & Grimmebergen, G. (2016):Grätensandsteine und andere Geschiebe des oberen Lias (Toarcium) aus Norddeutschland [Upper Liassic sandstones with fish remains (so-called Grätensandsteine) and other Toarcian glacial erratics from northern Germany]. Geschiebekunde aktuell 32 (4): 121–141, 12 Abb
- Lehmann, U. (1966). Dimorphismus bei Ammoniten der Ahrensburger Lias-Geschiebe. Paläontologische Zeitschrift, 40(1-2), 26–55.
- Hoffmann, R., Schultz, J. A., Schellhorn, R., Rybacki, E., Keupp, H., Lemanis, R., & Zachow, S. (2014). Non-invasive imaging methods applied to neo-and paleo-ontological cephalopod research.
- LürnG, G. (1995). Geschiebezählungen-eine terminologische Richtigstellung. Geschiebekunde aktuell, 11(4), 109–112.
- Hoffmann, K., & Martin, G. P. (1960). Die Zone desDactylioceras tenuicostatum (Toarcien, Lias) in NW-und SW-Deutschland. Paläontologische Zeitschrift, 34(2), 103–149.
- Schlögl, J., Košt’ák, M., & Hyžný, M. (2012). First record of a gladius-bearing coleoid Teudopsis bollensis Voltz (Cephalopoda, Coleoidea) in the Toarcian of the Western Carpathians (Slovakia). Paläontologische Zeitschrift, 86(4), 367–375.
- Riegraf, W. (1997). On the proposed conservation of the names Geopeltis Regteren Altena, 1949, Geoteuthis Muenster, 1843, Jeletzkyteuthis Doyle, 1990, Loligosepia Quenstedt, 1839, Parabelopeltis Naef, 1921, Paraplesioteuthis Naef, 1921 and Belemnotheutis montefiorei Buckman, 1880 (Mollusca, Coleoidea). BULLETIN OF ZOOLOGICAL NOMENCLATURE, 54, 184-184.
- Fuchs, D., & Weis, R. (2008). Taxonomy, morphology and phylogeny of Lower Jurassic loligosepiid coleoids (Cephalopoda). Neues Jahrbuch Für Geologie Und Paläontologie - Abhandlungen, 249(1), 93–112. doi:10.1127/0077-7749/2008/0249-0093
- Mikrofaunen aus Jura und Kreide insbesondere Nordwestdeutschlands. 1. Teil. Lias a—s. Abh. Preuss. Geol. L.A.N.F.I. 193, 1938.
- Kutscher, M. (1988). Zur Invertebratenfauna und Stratigraphie des oberen Pliensbachien von Grimmen (DDR), Echinodermata. Freiberger Forschungshefte C, 419, 62–70.
- Eichenberg, W. (1935). Holothurien-Kalkkörperchen aus dem Jura Norddeutschlands. Zeitschrift der Deutschen Geologischen Gesellschaft, 318-320.
- Bartenstein, H. (1936). Kalk-Körper von Holothurien in norddeutschen Lias-Schichten. Senckenbergiana, 18(1/2), 1–10.
- Frentzen, K. (1964). Funde von Holothurien-Kalkkörperchen im Jura des Oberrheingebietes. Beiträge zur naturkundlichen Forschung in Südwestdeutschland, 23, 31-51.
- Geinitz, F. E. (1884). Ueber die Fauna des Dobbertiner Lias. Zeitschrift der deutschen geologischen Gesellschaft, 566–583.
- Ansorge, J. (2007). Lower Jurassic clay pit of Klein Lehmhagen near Grimmen. The Central European Basin System–from the Bottom to the Top. Geo-Pomerania, Szczechin, 37–41.
- Audo, D., Schweigert, G., & Charbonnier, S. (2019, November). Proeryon, a geographically and stratigraphically widespread genus of polychelidan lobsters. In Annales de Paléontologie (p. 102376). Elsevier Masson.
- Leonowicz, P. (2011). Sedimentation of Lower Toarcian (Lower Jurassic) brackish deposits from the Częstochowa-Wieluń region (SW Poland). Acta Geologica Polonica, 61(2), 215–241.
- E. Herrig Zur Taxonomie und Evolution der Gattung Kinkelinella Martin, 1960 (Ostracoda) im Unteren Jura von Mittel- und Nordwesteuropa Zeitschrift für Geologische Wissenschaften, Berlin, 13 (6) (1985), pp. 715–723
- E. Herrig Die polycopiden Ostrakoden aus dem thüringischen Lias Zeitschrift für Geologische Wissenschaften, Berlin, 9 (6) (1981), pp. 675–696
- T. Pietrzenuk Zur Mikrofauna einiger Lias vorkommen in der Deutchen Demokratischen Republik Freiberger Forschunghelfe, H(C), 113 (1961), pp. 1–129
- E. Dreyer Mikrofossilem des Rät und Lias von SW-Brandenburg Geologische Jahrbuch, Hannover, 1 (1967), pp. 491–531
- E. Herrig Die Gattung Ogmoconchella—Arten (Ostracoda) im Lias von Thüringen Zeitschrift für Geologische Wissenschaften, Berlin, 9 (5) (1981), pp. 561–579
- E. Herrig Die Gattung Ogmoconcha Triebel, 1941 (Ostracoda) im Lias von Thüringen Zeitschrift für Geologische Wissenschaften, Berlin, 9 (2) (1981), pp. 207–219
- E. Herrig Ostrakoden der Gattungen Ledahia and Pseudohealdia (Familie: Healdiidae Harlton) aus dem Lias von Thüringen Zeitschrift für Geologische Wissenschaften, Berlin, 8 (12) (1980), pp. 1539–1551
- E. Herrig Ostracoden aus dem Ober-Domèrien von Grimmen westlich Griefswald (Teil I) Geologie, Berlin, 18 (1969), pp. 446–471
- Close E. Herrig, H. Richter Zur Entwicklung der Cytheropterinae (Ostracoda Crustacea) im oberen Lias von Mitteleuropa Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, Stuttgart, 180 (2) (1990), pp. 239-257
- E. Herrig Ostrakoden aus dem Lias von Thüringen. Die Familien Progonocytheridae, Cytherettidae und Brachycytheridae Zeitschrift für Geologische Wissenschaften, 10 (1982), pp. 1449–1461
- E. Herrig Ostracoden aus dem Lias von Thüringen. Die Familien Trachyleberididae, Paradoxostomatidae, Cytherellidae sowie Nachtrag zu den Paracyprididae Zeitschrift für Geologische Wissenschaften, 10 (1982), pp. 231–243
- E. Herrig Die Gattung Bairdia (Ostracoda, Crustacea) im Lias von Thüringen Zeitschrift für Geologische Wissenschaften, Berlin, 7 (5) (1979), pp. 641–661
- E. Herrig Ostrakoden aus dem Lias von Thüringen. Die Gattungen Bairdia (Teil II), Fabalacypris and Bairdiacypris Zeitschrift für Geologische Wissenschaften, Berlin, 7 (6) (1979), pp. 763–782
- Selden, Paul A.; Dunlop, Jason A. (2014). "The first fossil spider (Araneae: Palpimanoidea) from the Lower Jurassic (Grimmen, Germany)". Zootaxa. 3894 (1): 161–168. doi:10.11646/zootaxa.3894.1.13.
- Ansorge, J. (2003). Insects from the lower Toarcian of middle Europe and England. Acta zoologica cracoviensia, 46(SUPPL.), 291–310.
- Q. Q. Zhang, W. Mey, J. Ansorge, T. A. Starkey, L. T. McDonald, M. E. McNamara, E. A. Jarzembowski, W. Wichard, R. Kelly, X. Y. Ren, J. Chen, H. C. Zhang, and B. Wang. 2018. Fossil scales illuminate the early evolution of lepidopterans and structural colors. Science Advances 4:e1700988
- A. Handlirsch. 1906. Die Fossilen Insekten und die Phylogenie der Rezenten Formen, parts I–IV. Ein Handbuch fur Palaontologen und Zoologen 1–640
- W. Zessin. 1983. Revision der mesozoischen Familie Locustopsidae unter Berücksichtigung neuer Funde (Orthopteroida, Caelifera). Deutsche Entomologische Zeitschrift 30:173–237
- W. Zessin. 1987. Variabilität, Merkmalswandel und Phylogenie der Elcanidae im Jungpaläozoikum und Mesozoikum und die Phylogenie der Ensifera. Deutsche Entomologische Zeitschrift 34(1-3):1–76
- A. Handlirsch. 1939. Neue Untersuchungen über die fossilen Insekten mit Ergänzungen und Nachträgen sowie Ausblicken auf phylogenetische, palaeogeographische und allgemein biologische Probleme. II Teil. Annalen des Naturhistorischen Museums in Wien 49:1–240 [M. Clapham/J. Karr/M. Clapham]
- W. Zessin. 1987. Variabilität, Merkmalswandel und Phylogenie der Elcanidae im Jungpaläozoikum und Mesozoikum und die Phylogenie der Ensifera. Deutsche Entomologische Zeitschrift 34(1-3):1–76
- W. Zessin. 1988. Neue Saltatoria (Insecta) aus dem Oberlias Mitteleuropas. Freiberger Forschungshefte C 419:107–121
- J. Ansorge. 1996. Insekten aus dem oberen Lias von Grimmen (Vorpommern, Norddeutschland). Neue Paläontologische Abhandlungen 2:1–132
- J. Ansorge. 1999. Aenne liasina gen. et sp. n. - the most primitive non-biting midge (Diptera: Chironomidae: Aenninae subfam. n.) - from the Lower Jurassic of Germany. Polskie Pismo Entomologiczne 68:431–443
- Ansorge, J. (1993). Dobbertiniopteryx capniomimus gen. et sp. nov. die erste Steinfliege (Insecta: Plecoptera) aus dem europäischen Jura. Paläontologische Zeitschrift, 67(3-4), 287–292.
- W. Krzemiński and W. Zessin. 1990. The Lower Jurassic Limoniidae from Grimmen (GDR) (Dipt. Nematocera). Deutsche Entomologische Zeitschrift 37:39–43
- Kopeć, K., Ansorge, J., Soszyńska-Maj, A., & Krzemiński, W. (2018). Revision of the genus Mesotipula Handlirsch, 1920 (Diptera, Limoniidae, Architipulinae) from the Lower Jurassic of Northeast Germany. Historical Biology, 1–8.
- KRZEMIŃSKI, W., & ANSORGE, J. A new rhagionid fly from the Lower. Jurassic. Polskie Pismo Entomologiczne, 74, 3.
- A. Handlirsch. 1920. Palaeontologie. Handbuch der Entomologie 3:117–208
- N. S. Kalugina. 1985. Infraorders Psychodomorpha, Tipulomorpha and Culicomorpha. Dvukrylye nasekomye Yury Sibiri 33-113
- J. Ansorge. 1994. Tanyderidae and Psychodidae (Insecta: Diptera) from the Lower Jurassic of northeastern Germany. Palaeontologische Zeitschrift 68:199–210
- E. Lukashevich, J. Ansorge, W. Krzemiński and E. Krzemińska. 1998. Revision of Eoptychopterinae (Diptera: Eoptychopteridae). Polskie Pismo Entomologiczne 67:311-343
- ANSORGE, J. (2001). Lower Jurassic Hennigmatidae (Diptera) from Germany. Studia dipterologica, 8(1), 97–102.
- Ansorge, J. (1993). Parabittacus analis Handlirsch 1939 und Parabittacus lingula (Bode 1953), Neorthophlebiiden (Insecta: Mecoptera) aus dem Oberen Lias von Deutschland. Paläontologische Zeitschrift, 67(3–4), 293-298.
- A. Wendt. 1940. Liasocoris hainmülleri n. sp., eine fossile Wanze aus Mecklenburg. Archiv des Vereins der Freunde der Naturgeschichte in Mecklenburg, Neue Folge 15:18–20
- A. Handlirsch. 1921. Palaeontologie. Handbuch der Entomologie 3:209-304
- Y. A. Popov. 1992. Jurassic bugs (Hemiptera: Heteroptera) from the Museum of Natural History in Vienna. Annalen des Naturhistorischen Museums in Wien, Serie A 94:7–14
- D. E. Shcherbakov. 2012. More on Mesozoic Membracoidea (Homoptera). Russian Entomological Journal 21:15–22
- G. Enderlein. 1909. Zur Kenntnis frühjurassischer Copeognathen und Coniopterygiden und über das Schicksal der Archipsyiliden. Zoologischer Anzeiger 34:770-776
- Ansorge, J., & Schlüter, T. (1990). The earliest chrysopid: Liassochrysa stigmatica ng, n. sp. from the Lower Jurassic of Dobbertin, Germany. Neuroptera International, 6(2), 87-93.
- Krüger, L. (1922). Hemerobiidae. Beiträge zu einer Monographie der Neuropteren-Familie der Hemerobiiden. Stettiner Entomologische Zeitung, 83, 138-172.
- A. Bode. 1953. Die Insektenfauna des Ostniedersachsischen Oberen Lias. Palaeontographica Abteilung A 103:1-375
- W. Zessin. 1982. Durchsicht einiger liassischer Odonatopteroida unter Berücksichtigung neuer Funde von Dobbertin in Mecklenburg. Deutsche Entomologische Zeitschrift 29:101-106
- Zessin, W., & Ansorge, J. (1987). Magnasupplephlebia intercalaria n. sp.‐eine neue Anisozygopterenart aus dem oberen Lias von Mitteleuropa.(Insecta, Odonata). Deutsche Entomologische Zeitschrift, 34(4‐5), 383-386.
- J. Ansorge. 2004. Insekten aus Liasgeoden der Ahrensburger Geschiebesippe - mit einem Ausblick auf lokale Anreicherungen von Liäsgeoden in Mecklenburg-Vorpommern. Archiv für Geschiebekunde 3:779–784
- J. Cowley. 1942. Descriptions of some genera of fossil Odonata. Proceedings of the Royal Entomological Society of London. Series B, Taxonomy 11:63–78
- A. P. Rasnitsyn, J. Ansorge, and H. C. Zhang. 2006. Ancestry of the orussoid wasps, with description of three new genera and species of Karatavitidae (Hymenoptera = Vespida: Karatavitoidea stat. nov.). Insect Systematics & Evolution 37:179–190
- W. Zessin. 1981. Ein Hymenopterenflegel aus dem oberen Lias bei Dobbertin, Bezirk Schwerin. Zeitschrift für Geologische Wissenschaften 9:713–717
- A. P. Rasnitsyn, J. Ansorge, and W. Zessin. 2003. New hymenopterous insects (Insecta: Hymenoptera) from the lower Toarcian (Lower Jurassic) of Germany. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 227:321-342
- P. Vršanský and J. Ansorge. 2007. Lower Jurassic cockroaches (Insecta: Blattaria ) from Germany and England. African Invertebrates 48(1):103-126
- F. E. Geinitz. 1883. Die Flötzformationen Mecklenburgs. Archiv des Vereins der Freunde der Naturgeschichte in Mecklenburg 37:1–151
- S. H. Scudder. 1886. A review of Mesozoic cockroaches. Memoirs of the Boston Society of Natural History 3:439-484
- E. Geinitz. 1894. Die Kanferreste des Dobbertiner Lias. Archiv des Vereins der Freunde der Naturgeschichte in Mecklenburg 48:71–78
- E. E. Maxwell and S. Stumpf. 2017. Revision of Saurorhynchus (Actinopterygii: Saurichthyidae) from the Early Jurassic of England and Germany. European Journal of Taxonomy 321:1–29
- Stumpf, S. (2017). A Synoptic Review of the Vertebrate Fauna from the "Green Series" (Toarcian) of Northeastern Germany with Descriptions of New Taxa: A Contribution to the Knowledge of Early Jurassic Vertebrate Palaeobiodiversity Patterns (Doctoral dissertation, Mathematisch-Naturwissenschaftliche Fakultät der Ernst-Moritz-Arndt-Universität Greifswald).
- J. J. Hornung and S. Sachs. 2020. First record of Gyrosteus mirabilis (Actinopterygii, Chondrosteidae) from the Toarcian (Lower Jurassic) of the Baltic region. PeerJ 8:e8400
- Konwert, M., & Hörnig, M. (2018). Grimmenichthys ansorgei, gen. et sp. nov. (Teleostei, “Pholidophoriformes”), and other “pholidophoriform” fishes from the early Toarcian of Grimmen (Mecklenburg-Western Pomerania, Germany). Journal of Vertebrate Paleontology, 38(3), e1451871. doi:10.1080/02724634.2018.1451871
- Agassiz, L. (1844). Tableau général des poissons fossiles rangés par terrains. Imprimerie de Petitpierre.
- Lehmann, U. (1971). Faziesanalyse der Ahrensburger Liasknollen auf Grund ihrer Wirbeltierreste. Mitteilungen aus dem Geologischen Institut der Technischen Universität Hannover, 10, 21-42.
- Konwert, M., and S. Stumpf. 2017. Exceptionally preserved Leptolepidae (Actinopterygii, Teleostei) from the late Early Jurassic Fossil Lagerstatten of Grimmen and Dobbertin (Mecklenburg-Western Pomerania, Germany). Zootaxa 4243:249–296
- Agassiz, L. (1832) Untersuchungen über die fossilen Fische der Lias-Formation—Aus einem Briefe des Vfs. an Professor Bronn. Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 3, 139–149.
- Thies, D. 1989. Sinneslinien bei dem Knochenfisch Lepidotes elvensis (Blainville 1818) (Actinopterygii, Semionotiformes) aus dem Oberlias (Unter-Toarcium) von Grimmen in der DDR. Neues Jahrbuch fur Geologie und Palaontologie, Monatshefte 1989:692–704.
- Thies, D., Stevens, K., & Stumpf, S. (2019). Stomach contents of the Early Jurassic fish† Lepidotes Agassiz, 1832 (Actinopterygii, Lepisosteiformes) and their palaeoecological implications. Historical Biology, 1-12.
- S. Stumpf, J. Ansorge, C. Pfaff and J. Kriwet. 2017. Early Jurassic diversification of pycnodontiform fishes (Actinopterygii, Neopterygii) after the end-Triassic extinction event: evidence from a new genus and species, Grimmenodon aureum. Journal of Vertebrate Paleontology 37:e1344679
- Thies, D. (1988). Dapedium pholidotum (AGASSIZ, 1832) - (Pisces, Actinopterygii) from Unter-Toarcium NW-Germany. Geologica et Palaeontologica , (22), 89–121.
- Böhme, M., Ilg, A. 2003. fosFARbase, http://www.wahre-staerke.com.
- Kovalchuk, О. М., & Anfimova, G. V. (2020). Lepisosteiform fish (Holostei) ganoid scales from the middle jurassic deposits of Ukraine. Zoodiversity, 54(1), 35-42.
- M. W. Maisch and J. Ansorge. 2004. The Liassic ichthyosaur Stenopterygius cf. quadriscissus from the Lower Toarcian of Dobbertin (northeastern Germany) and some considerations on Lower Toarcian marine reptile palaeobiogeography. Palaeontologische Zeitschrift 78(1):161–171
- GEINITZ, F.E. 1900a. Ichthyosaurus von Dobbertin.- Archiv des Vereins der Freunde der Naturgeschichte in Mecklenburg 54: 382-383.
- ZESSIN, W. 1995. Saurierfund im Lias epsilon von Grimmen, Kreis Nordvorpommern (Fundbericht). - Geschiebkunde Aktuell 11 (4): 113.
- ZESSIN, W. 1998.20 Jahre geowissenschaftliche Freizeitforschung in Westmecklenburg.- Geschiebekunde Aktuel114 (1): 1-10
- ZESS1N, W. 2001. Ichthyo-Saurierfunde und KrokodilschSxtel aus dem Lias von Klein Lehmhagen bei Grimmen, Kreis Nordvorpommern. - NABU Nachrichten Mecklenburg-Vorpommern 2001 (1): 7-9.
- LEHMANN, U. 1968. Stratigraphie und Ammonitenfiihrung der Ahrensburger Glazial-Geschiebe aus dem Lias epsilon (= Unt. Toarcium). - Mitteilung des Geologischen Staatsinstituts Hamburg 37: 41-68.
- LIERL, H.-J. 1990. Die Ahrensburger Geschiebesippe. - Fossilien 1990 (6): 256-267.
- MOTHS, H. 1994. Ein seltenes Ichthyosaurus-Gebil3 als Geschiebe aus Mecklenburg. - Der Geschiebesammler 27 (1): 15-22.
- Sachs, S., Hornung, J. J., Lierl, H.-J., & Kear, B. P. (2016). Plesiosaurian fossils from Baltic glacial erratics: evidence of Early Jurassic marine amniotes from the southwestern margin of Fennoscandia. Geological Society, London, Special Publications, 434(1), 149–163. doi:10.1144/sp434.14
- WINCIERZ, J. 1967. Ein Steneosaurus-Fund aus dem nordwestdeutschen Oberen Lias. Paläontologische Zeitschrift 41:60–72.
- STUMPF, S. 2016. New information on the marine reptile fauna from the lower Toarcian (Early Jurassic) “Green Series” of North-Eastern Germany. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 280(1):87–105.
- Mueller-Töwe, I. J. (2006). Feeding options in Steneosaurus bollensis (Mesoeucrocodylia. Thalattosuchia). Hantkeniana Spec, 5, 46–48.
- F. v. Huene. 1966. Ein Megalosauriden-Wirbel des Lias aus norddeutschem Geschiebe [A megalosaurid vertebra from the Lias of a north German boulder]. Neues Jahrbuch für Geologie und Paläontologie 1966(5):318-319
- Stumpf, S., & Meng, S. (2013). Dinosaurier aus Nordostdeutschland: Verschleppt. Biologie in unserer Zeit, 43(6), 362-368.
- S. Stumpf (2013): Die unterjurassische Wirbeltierfauna der Tongrube Grimmen (Nordostdeutschland). gfg-vorpommern:https://www.gfg-vorpommern.de/archive/843?pdf=843
- S. Stumpf, J. Ansorge, and W. Krempien. 2015. Gravisaurian sauropod remains from the marine late Early Jurassic (Lower Toarcian) of North-Eastern Germany. Geobios 48:271-279
- Haubold, H. 1990. Ein neuer Dinosaurier (Ornithischia, Thyreophora) aus dem Unteren Jura des nördlichen Mitteleuropa. Revue de Paleobiologie 9(1):149-177. [In German]
- Haubold, H. (1991). Der Greifswalder Dinosaurier ‘‘Emausaurus’’. Fundgrube, 27(2), 51-60.
- LILPOP J. 1917. Mikroskopisch-anatomische Untersuchungen der Mineralkohlen. Bull. Acad. Sci. Crac.,Sci. Math. Nat., Ser. B: Sci. Nat., 1917: 6–24
- A. Teofilak Liassic heavy minerals and their origin Bulletin de l’Academie Polonaise des Sciences, Séries Sciences Géologie et Géographie, 14 (1966), pp. 17–21
- Rybicki, M., Marynowski, L., Misz-Kennan, M., & Simoneit, B. R. T. (2016). Molecular tracers preserved in Lower Jurassic 'Blanowice brown coals' from southern Poland at the onset of coalification: Organic geochemical and petrological characteristics. Organic Geochemistry, 102, 77–92. doi:10.1016/j.orggeochem.2016.09.012
- RYBICKI, M., MARYNOWSKI, L., & SIMONEIT, B. R. (2015): Sesquiterpenoids and diterpenoids from Lower Jurassic sub-bituminous coal from the “Blanowice Formation”, southern Poland. In XXII nd Meeting of the Petrology Group of the Mineralogical Society of Poland (p. 87).
- Rybicki, M., Marynowski, L., & Simoneit, B. R. (2017). Benzohopane Series, Their Novel Di-, Tri-, and Tetraaromatic Derivatives, and Diaromatic 23-and 24-Norbenzohopanes from the Lower Jurassic Blanowice Formation, Southern Poland. Energy & Fuels, 31(3), 2617–2624.
- Pointer, R. (2019). Fire & Global Change During Key Intervals of the Late Triassic & Early Jurassic with a Focus on the Central Polish Basin.
- Marcinkiewicz, T., Fijałkowska-Mader, A., & Pieńkowski, G. (2014). Megaspore zones of the epicontinental Triassic and Jurassic deposits in Poland–overview. Biuletyn Państwowego. Instytutu Geologicznego, 457, 15–42.
- DOMAGAŁA M. & KOŁCON I. 1983. Zbiorowiska roślinności węglotwórczej liasowego węgla brunatnegoz Poręby koło Zawiercia (summary: Vegetation assemblages forming Lias brown coals in the Poręba area near Zawiercie). Kwart. Geol., 27(3): 503–516.
- Grabowska,I. et al. (1970) Flora from the Lower and Middle Jurassic microflora Geology of Poland. Catalogue of Fossils. Mesozoic Vol. 2 # 2 p. 47–53
- Marcinkiewicz, T. (1973). Otozamites falsus (Bennettitales) from the Upper Liassic of the Holy Cross Mts, Poland. Acta Palaeontologica Polonica, 18(2).
- Dadlez, R., Kopik, J., Marcinkiewicz, T., & Szymborsky, L. (1964). Results obtained in Bore-Hole Mechowo IG 1. Inst. Geol. Bulletin, 189, 1-156.
- Rogalska,M. (1971) Division of the Liassic deposits in Poland (Except for the Carpathian area) based on microscope examinations. M©moires du Bureau des Recherches G©ologiques et MiniЁres (BRGM) Vol. 75 P. 201- 210
- Marcinkiewicz, T. (1960). Megaspore analysis of Jurassic sediments near Gorzow Slaski Praszha (Cracow-Wielun region). Kwart. geol, 4, 713–33.
- Malinowska, L. (1980). Atlas skamieniałości przewodnich i charakterystycznych. Część 2b, mezozoik/jura. Budowa Geologiczna Polski, 3.
- Marcinkiewicz, T. (1974). Występowanie megaspor w obrębie zaburzonych warstw i liasu w profilu Koszalina. Geological Quarterly, 18(3), 595–601.
- Marcinkiewicz,T. et al. (1960) Age of the Upper Helenow Beds, (Lias), in view of mega and microspore investigations, (Geological section Gorzow Slaski-Praska). [ Wiek Warstw Helenowskich Gornych (Lias) w Przekroju Geologicznym Gorzow Slaski -Praszka w Swietle badan Mega-i Mikrsporowych. ] Kwartalnik Geologiczny (Instytut Geologiczy) Vol. 4 # 2 pp. 386–398
- MARCINKIEWICZ, T., FIJAŁKOWSKA-MADER, A. N. N. A., & PIEŃKOWSKI, G. (2014). Poziomy megasporowe epikontynentalnych utworów triasu i jury w Polsce–podsumowanie. Biuletyn Państwowego Instytutu Geologicznego, 457, 15–42.
- Kovach, W. L., & Dilcher, D. L. (1985). Morphology, ultrastructure, and paleoecology of Paxillitriletes vittatus sp. nov. from the mid‐Cretaceous (Cenomanian) of Kansas. Palynology, 9(1), 85-94.
- Marcinkiewicz, T. E. R. E. S. A. (1989). Remarks on agglomerations of megaspores Minerisporites institus Marc. Acta Palaeobotanica, 29, 221–224.
- Dyakowska,J. (1958) The angiospermoid pollen from the Liassic flora in Poland Veroffentlichungen des Geobotanischen Inst.Rubel i N Zurich,Verhandlu,4th Int.Meet,Quat.Botan. Vol. 34 pp. 42– 43
- Hemsley, A. R., Collinson, M. E., Kovach, W. L., Vincent, B., & Williams, T. (1994). The role of self-assembly in biological systems: evidence from iridescent colloidal sporopollenin in Selaginella megaspore walls. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 345(1312), 163-173.
- Ziaja, J. (2006). Lower Jurassic spores and pollen grains from Odrowąż, Mesozoic margin of the Holy Cross Mountains, Poland. Acta Palaeobotanica, 46(1), 3-83.
- Van Konijnenburg-van Cittert, J. H. (1992). An enigmatic Liassic microsporophyll, yielding Ephedripites pollen. Review of palaeobotany and palynology, 71(1-4), 239-254.
- Tekleva, M. V., Krassilov, V. A., Kvacek, J., & Van Konijnenburg-van Cittert, J. H. A. (2006). Pollen genus Eucommiidites: ultrastructure and affinities. ACTA PALAEOBOTANICA-KRAKOW-, 46(2), 137.
- Crane, P. R. (1996). The fossil history of the Gnetales. International Journal of Plant Sciences, 157(S6), S50-S57.
- Rydin, C., Pedersen, K. R., Crane, P. R., & Friis, E. M. (2006). Former diversity of Ephedra (Gnetales): evidence from early Cretaceous seeds from Portugal and North America. Annals of Botany, 98(1), 123-140.
- Philippe, M., & Thevenard, F. (1996). Distribution and palaeoecology of the Mesozoic wood genus Xenoxylon: palaeoclimatological implications for the Jurassic of Western Europe. Review of Palaeobotany and Palynology, 91(1-4), 353–370.
- Philippe, M., Thévenard, F., Nosova, N., Kim, K., & Naugolnykh, S. (2013). Systematics of a palaeoecologically significant boreal Mesozoic fossil wood genus, Xenoxylon Gothan. Review of Palaeobotany and Palynology, 193, 128-140.
- Migier T. 1978. Nowe stanowiska flory jurajskiej w Lubelskim Zagłębiu Węglowym. Materiały III Naukowej Konferencji Paleontologów poświęconej badaniom regionu górnośląskiego oraz karbonu LZW i GZW. Streszczenia komunikatów: 33–34.Uniwersytet Śląski, Katowice.
- Ruebsam, W., Pieńkowski, G., & Schwark, L. (2020). Toarcian climate and carbon cycle perturbations–its impact on sea-level changes, enhanced mobilization and oxidation of fossil organic matter. Earth and Planetary Science Letters, 546, 116417.
- Samsonowicz, J. (1929). Cechsztyn, trias i lias na północnem zboczu Łysogór.. PIG, Sprawozdania,t. V, z. 1-2
- Szydeł, Z., & Szydeł, R. (1981). Profil utworów liasu na obszarze Lubelskiego Zagłębia Węglowego. Przegląd Geologiczny, 29(11), 568-571.
- Ernst, W. (1987). Lias-Tontagebau Grimmen. In Exkursionsführer zum Otto-Jaekel-Symposium in Greifswald (5.–8. 10. 1987). Xerocopy Univ. Greifswald (pp. 17-31).
- FUCHS, A. & W. KINZEL (1991): Ein neues geologisches Naturdenkmal in Mecklenburg: "Lias von Dobbertin".- Geschiebekunde aktuell 7 (3): 133.
- FUCHS, A. & W. ZIMMERLE (1991): Zur Bedeutung des Lias-Aufschlusses von Dobbertin (Mecklenburg) – Unter Betrachtung vorläufiger tonmineralogischer Untersuchungen.-Geschiebekunde aktuell 7 (4): 179-186
- Pieńkowski, G., Hesselbo, S. P., Barbacka, M., & Leng, M. J. (2020). Non-marine carbon-isotope stratigraphy of the Triassic-Jurassic transition in the Polish Basin and its relationships to organic carbon preservation, pCO2 and palaeotemperature. Earth-Science Reviews, 103383.
- Kozydra, Z. (1960). Zarys wykształcenia litologicznego serii “ciechocińskiej” liasu w rejonie Przysuchej. Przegląd Geologiczny, 8(9), 456.
- Pacyna, G. (2013). Critical review of research on the Lower Jurassic flora of Poland. Acta Palaeobotanica, 53(2), 141-163.