Archosauriformes
Archosauriformes (Greek for 'ruling lizards', and Latin for 'form') is a clade of diapsid reptiles that developed from archosauromorph ancestors some time in the Late Permian (roughly 250 million years ago). It was defined by Jacques Gauthier (1994) as the clade stemming from the last common ancestor of Proterosuchidae and Archosauria (the group that contains crocodiles, pterosaurs and dinosaurs (including birds);[3] Phil Senter (2005) defined it as the most exclusive clade containing Proterosuchus and Archosauria.[4]
| Archosauriformes | |
|---|---|
 | |
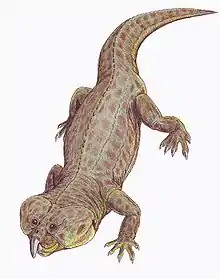
| Life restoration of a Proterosuchus fergusi | |
 | |
| American alligator (A. mississippiensis) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Crocopoda |
| Clade: | Archosauriformes Gauthier, 1986 |
| Subgroups[1] | |
| |
These reptiles, which include members of the family Proterosuchidae and more advanced forms, were originally superficially crocodile-like animals with sprawling gaits and long snouts. Unlike the bulk of their therapsid contemporaries, the proterosuchids survived the catastrophe at the end of the Permian. Within a few million years after the beginning of the Triassic, the archosauriformes had diversified past the "proterosuchian" grade. Other archosauriforms include the Erythrosuchidae (some of the earliest sauropsid apex predators), the Euparkeriidae (small, agile reptiles), and a variety of other strange reptiles such as proterochampsids, Vancleavea, Doswellia, and Asperoris. The most successful archosauriforms, and the only members to survive into the Jurassic, were the archosaurs. Archosauria includes crocodylians, birds, and extinct relatives such as aetosaurs, rauisuchids, pterosaurs, and non-avian dinosaurs.[5]
Pre-Euparkeria Archosauriformes have previously been included in the suborder Proterosuchia of the order Thecodontia. Under modern cladistic methodology, Proterosuchia and Thecodontia have been rejected as paraphyletic assemblages, and the pre-archosaurian taxa are simply referred to as basal Archosauriformes. In 2016, Martin Ezcurra provided a name for the clade including all archosauriforms more crownward than erythrosuchids. He named the clade Eucrocopoda, which translates to "true crocodile feet" in reference to the possession of a crocodilian-style crurotarsal ankle.[1]
Metabolism
Vascular density and osteocyte density, shape and area have been used to estimate the bone growth rate of archosaurs, leading to the conclusion that this rate had a tendency to grow in ornithodirans and decrease in pseudosuchians.[6] The same method also supports the existence of high resting metabolical rates similar to those of living endotherms (mammals and birds) in the Prolacerta-Archosauriformes clade that were retained by most subgroups, though decreased in Proterosuchus, Phytosauria and Crocodilia.[7] Erythrosuchids and Euparkeria are basal archosauriforms showing signs of high growth rates and elevated metabolism, with Erythrosuchus possessing a rate similar of the fastest-growing dinosaurs. Sexual maturity in those Triassic taxa was probably reached quickly, providing advantage in a habitat with unpredictable variation from heavy rainfall to drought and high mortality. Vancleavea and Euparkeria, which show slower growth rates compared to Erythrosuchus, lived after the climatic stabilization. Early crown archosaurs possessed increased growth rates, which were retained by ornithodirans.[8] Ornithosuchians and poposaurs are stem-crocodilians that show high growth rates similar to those of basal archosauriforms.[9]
Developmental, physiological, anatomical and palaeontological lines of evidence indicate that crocodilians evolved from endothermic ancestors. Living crocodilians are ambush predators adapted to a semi-aquatic lifestyle that benefits from ectothermy due to the lower oxygen intake that allows longer diving time. The mixing of oxygenated and deoxygenated blood in their circulatory system is apparently an innovation that benefits ectothermic life. Earlier archosaurs likely lacked those adaptations and instead had completely separated blood as birds and mammals do.[10][11] A similar process occurred in phytosaurs, which were also semi-aquatic.[12]
The similarities between pterosaur, ornithischian and coelurosaurian integument suggest a common origin of thermal insulation (feathers) in ornithodirans at least 250 million years ago.[13][14] Erythrosuchids living in high latitudes might have benefited from some sort of insulation.[12] If Longisquama was an archosauromorph, it could be associated with the origin of feathers.[15][12]
Relationships
Below is a cladogram from Nesbitt (2011):[16]
| Archosauriformes |
*Note: Phytosaurs were previously placed within Pseudosuchia, or crocodile-line archosaurs. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Below is a cladogram from Sengupta et al. (2017),[17] based on an updated version of Ezcurra (2016)[1] that reexamined all historical members of the "Proterosuchia" (a polyphyletic historical group including proterosuchids and erythrosuchids). The placement of fragmentary taxa that had to be removed to increase tree resolution are indicated by dashed lines (in the most derived position that they can be confidently assigned to). Taxa that are nomina dubia are indicated by the note "dubium". Bold terminal taxa are collapsed.[1]
| Crocopoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sources
- Gauthier, J. A. (1986). "Saurischian monophyly and the origin of birds". In Padian, K. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences. 8. California Academy of Sciences. pp. 1–55. ISBN 978-0-940228-14-6.
- Gauthier, J. A.; Kluge, A. G.; Rowe, T. (June 1988). "Amniote phylogeny and the importance of fossils" (PDF). Cladistics. John Wiley & Sons. 4 (2): 105–209. doi:10.1111/j.1096-0031.1988.tb00514.x. hdl:2027.42/73857. S2CID 83502693.
References
- Ezcurra, Martín D. (2016-04-28). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. ISSN 2167-8359. PMC 4860341. PMID 27162705.
- Sookias, R. B.; Sullivan, C.; Liu, J.; Butler, R. J. (2014). "Systematics of putative euparkeriids (Diapsida: Archosauriformes) from the Triassic of China". PeerJ. 2: e658. doi:10.7717/peerj.658. PMC 4250070. PMID 25469319.
- Gauthier J. A. (1994): The diversification of the amniotes. In: D. R. Prothero and R. M. Schoch (ed.) Major Features of Vertebrate Evolution: 129-159. Knoxville, Tennessee: The Paleontological Society.
- Phil Senter (2005). "Phylogenetic taxonomy and the names of the major archosaurian (Reptilia) clades". PaleoBios. 25 (2): 1–7.
- Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin
- Cubo, Jorge; Roy, Nathalie Le; Martinez-Maza, Cayetana; Montes, Laetitia (2012). "Paleohistological estimation of bone growth rate in extinct archosaurs". Paleobiology. 38 (2): 335–349. doi:10.1666/08093.1. ISSN 0094-8373. S2CID 84303773.
- Legendre, Lucas J.; Guénard, Guillaume; Botha-Brink, Jennifer; Cubo, Jorge (2016-11-01). "Palaeohistological evidence for ancestral high metabolic rate in archosaurs". Systematic Biology. 65 (6): 989–996. doi:10.1093/sysbio/syw033. ISSN 1063-5157. PMID 27073251.
- Botha-Brink, Jennifer; Smith, Roger M. H. (2011-11-01). "Osteohistology of the Triassic archosauromorphs Prolacerta, Proterosuchus, Euparkeria, and Erythrosuchus from the Karoo Basin of South Africa". Journal of Vertebrate Paleontology. 31 (6): 1238–1254. doi:10.1080/02724634.2011.621797. ISSN 0272-4634. S2CID 130744235.
- de Ricqlès, Armand; Padian, Kevin; Knoll, Fabien; Horner, John R. (2008-04-01). "On the origin of high growth rates in archosaurs and their ancient relatives: Complementary histological studies on Triassic archosauriforms and the problem of a "phylogenetic signal" in bone histology". Annales de Paléontologie. 94 (2): 57–76. doi:10.1016/j.annpal.2008.03.002. ISSN 0753-3969.
- Seymour, Roger S.; Bennett‐Stamper, Christina L.; Johnston, Sonya D.; Carrier, David R.; Grigg, Gordon C. (2004-11-01). "Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution" (PDF). Physiological and Biochemical Zoology. 77 (6): 1051–1067. doi:10.1086/422766. hdl:2440/1933. ISSN 1522-2152. PMID 15674775. S2CID 10111065.
- Summers, Adam P. (April 2005). "Warm-hearted crocs". Nature. 434 (7035): 833–834. Bibcode:2005Natur.434..833S. doi:10.1038/434833a. ISSN 1476-4687. PMID 15829945. S2CID 4399224.
- "Dinosaur Renaissance". Scientific American. Retrieved 2020-05-03.
- Yang, Zixiao; Jiang, Baoyu; McNamara, Maria E.; Kearns, Stuart L.; Pittman, Michael; Kaye, Thomas G.; Orr, Patrick J.; Xu, Xing; Benton, Michael J. (January 2019). "Pterosaur integumentary structures with complex feather-like branching". Nature Ecology & Evolution. 3 (1): 24–30. doi:10.1038/s41559-018-0728-7. ISSN 2397-334X. PMID 30568282. S2CID 56480710.
- Benton, Michael J.; Dhouailly, Danielle; Jiang, Baoyu; McNamara, Maria (2019-09-01). "The early origin of feathers". Trends in Ecology & Evolution. 34 (9): 856–869. doi:10.1016/j.tree.2019.04.018. ISSN 0169-5347. PMID 31164250.
- Buchwitz, Michael; Voigt, Sebastian (2012-09-01). "The dorsal appendages of the Triassic reptile Longisquama insignis: reconsideration of a controversial integument type". Paläontologische Zeitschrift. 86 (3): 313–331. doi:10.1007/s12542-012-0135-3. ISSN 1867-6812. S2CID 84633512.
- Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades" (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112. S2CID 83493714.
- Sengupta, S.; Ezcurra, M.D.; Bandyopadhyay, S. (2017). "A new horned and long-necked herbivorous stem-archosaur from the Middle Triassic of India". Scientific Reports. 7 (1): 8366. doi:10.1038/s41598-017-08658-8. PMC 5567049. PMID 28827583.
External links
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||