Canardia
Canardia is an extinct genus of aralosaurin lambeosaurine dinosaur known from the Late Cretaceous Marnes d'Auzas Formation (late Maastrichtian stage) of Haute-Garonne department, in Occitanie region, southwestern France. The type species Canardia garonnensis was first described and named by Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete and Àngel Galobart in 2013.[1] The name of the genus comes from “canard”, the French word for “duck”, an allusion to the fact that this animal belongs to the hadrosaurids which are also known as duck-billed dinosaurs. The specific epithet garonnensis refers to the Haute-Garonne department where this dinosaur has been found. Canardia was one of the last non-avian dinosaurs and lived between 67,5 and 66 my on the former Ibero-Armorican Island, which included much of France and Spain.[1]
| Canardia | |
|---|---|
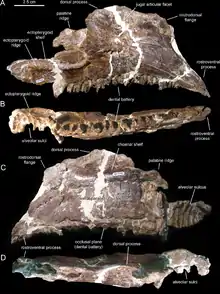 | |
| Holotype maxilla of Canardia garonnensis from the basal part of the Marnes d'Auzas Formation | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Suborder: | †Ornithopoda |
| Family: | †Hadrosauridae |
| Subfamily: | †Lambeosaurinae |
| Genus: | †Canardia Prieto-Márquez et al., 2013 |
| Type species | |
| †Canardia garonnensis Prieto-Márquez et al., 2013 | |
Discovery and naming

Canardia is known by various cranial and postcranial remains found in several sites in southwestern France (Haute-Garonne Department).[1] The type locality of Tricouté 3, near the village of Marignac-Laspeyres, was discovered in 1999 by a team of The Musée des Dinosaures d’Espéraza.[3] This site is located in the basal part of the Marnes d’Auzas Formation, which corresponds to a laguno-continental environment.[3][1] The hadrosaur remains were preserved in a small lenticular marly-sandstone level within a thick sandstone bed.[3][1] They belong to at least two different individuals as indicated by the presence of two left scapulae among the samples. The cranial elements include a nearly complete right maxilla (the holotype), a partial left maxilla, a right prefrontal, an incomplete right postorbital, a left quadrate, a right pterygoid, a partial left surangular, a right articular, an isolated dentary tooth crown, and a partial dental battery of the lower jaw. The postcranial material is represented by two partial left scapulae, an incomplete left sternal plate, a left humerus, and a partial right pubis. In 2003, these specimens were provisionally attributed to Pararhabdodon sp., a Spanish genus that at the time was the only known lambeosaurin in Europe. It was not until 2013 that Prieto-Márquez and colleagues recognized these bones as those of a new genus.[3][1] All bones are of relatively small sizes (the holotype maxilla is 16,9 cm, the scapula is 21,8 cm, and the humerus is only 19,4 cm) and belong to immature individuals.[3]
The same authors have also attributed to Canardia an associated maxilla and quadrate found in marine deposits of the Marly Limestone of Gensac Formation in Larcan quarry, about twenty km west of Marignac-Laspeyres.[1] The two bones, first described in 2010 by Bilotte et al., were located approximately 1 m under an iridium level marking the Cretaceous-Paleogene boundary.[2] This level is a lateral equivalent to the uppermost part of the Marne d’Auzas Formation. This shows that the Larcan individual is geologically younger than the Marignac-Laspeyres specimens. This discovery also indicates that Canardia was one of the last non-avian dinosaurs to exist in Europe just before the Cretaceous-Paleogene extinction event.[2][1] The Larcan specimen belongs to a juvenile individual (the length of the maxilla is 11,5 cm), whose remains were carried by a river to the sea.[2]
Description

The main feature of Canardia is the maxilla characterized by an enlarged rostrodorsal region that forms a prominent subrectangular flange that rises vertically above the rostroventral process. This character is also known in Aralosaurus tuberiferus from Kazakhstan that lived 20 million years before Canardia. However, the maxilla of Canardia differs from that of Aralosaurus in its subhorizontal ectopterygoid shelf (it is parallel to the posterior segment of the dental row) whereas it is clearly inclined in Aralosaurus. The maxilla of Canardia shows at least 26 teeth positions[1] against 30 in Aralosaurus,[4] but the entire tooth row is not preserved in the French form. The maxillary teeth have a single median carina while the dentary teeth have an accessory ridge in front of the sub-central carina, as is the case in Aralosaurus and in many other hadrosaurs.[1][4] Canardia differs also from Aralosaurus by the shape of the prefrontal in dorsal view. In Canardia, the rostroventral process of the prefrontal is rostrocaudally narrow, whereas in Aralosaurus, the same bone process is broad and laterally well-exposed. In addition, the prefrontal of Canardia has a dorsomedial flange that is absent in Aralosaurus.[1] Canardia and Aralosaurus are closely related, and Prieto-Márquez and colleagues brought them together in a new group of basal lambeosaurines which they call Aralosaurini.[1]
It is not known if Canardia had a hollow bony crest on the skull like most lambeosaurines. Its close relative Aralosaurus appears to have had a hollow structure in front of the orbits.[5] However, as only a fragment of this structure has been preserved, its size and shape are unfortunately unknown. Maybe Canardia had a similar structure, but the discovery of more complete cranial remains is necessary to make comparisons.
Phylogeny
With its sister group Aralosaurus tuberiferus, Canardia garonnensis form a small clade, the tribe of Aralosaurini. These two genera are the most basal Lambeosaurinae.
The phylogenetic analyzes carried out by Prieto-Márquez and his colleagues led to the following cladogram:[1]
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Palaeoecology
Paleobiogeography

The close relationship of Canardia with Aralosaurus confirms the existence of faunal exchange in the Late Cretaceous between the islands of the European archipelago and the Asian continent.[1][6] Canardia and Aralosaurus are temporally distant for at least 20 million years. As no lambeosaurines were known in the rich localities of late Campanian and early Maastrichtian ages of the Ibero-Armorican Island, it was suggested that the ancestors of Canardia had to reach this island rather late, perhaps at the end of the early Maastrichtian or during the late Maastrichtian.[1] The discovery later in Spain of numerous lambeosaurines remains in the highest levels of the lower Maastrichtian argues for the first hypothesis.[7][8] This migration was probably carried out in several stages, firstly by temporary terrestrial links between the landmass of Western Asia and the eastern islands of the European archipelago, then between the different European islands up to the Ibero-Armorican Island.[6] A similar scenario probably applies to the presence on this island during the late Maastrichtian of another lambeosaurine with Asian affinity, the genus Pararhabdodon which is close to the Chinese Tsintaosaurus (which lived 15 million years earlier). Both Canardia and Pararhabdodon belong to lambeosaurines groups that are rather archaic for their time (Aralosaurini and Tsintaosaurini respectively) and the Ibero-Armorican Island was perhaps an ultimate refuge for these ancient groups of lambeosaurines hadrosaurs.[1]
Palaeoenvironment
The Marnes d’Auzas Formation is 100 m thick. It corresponds to sediments whose depositional environment evolved from the paralic domain (coastal lagoons, tidal marsh, tidal muddy channel) at the base of the formation, towards a more continental domain (alluvial plain, fluvial channels) in its upper part.[3] From a paleogeographic point of view, the sites of the Marnes d’Auzas Formation were located on the west coast of the Ibero-Armorican Island (which included much of France and Spain), facing the Atlantic Gulf.[6]
Few other vertebrates fossils have been found with the Canardia remains. The type locality of Tricouté 3, in the basal part of the Marnes d’Auzas Formation, has yielded only a crocodilian tooth,[3] whereas the marine Marly limestone of Gensac Formation (the lateral equivalent of the top of the Marnes d’Auzas) has not yet yielded other terrestrial vertebrates remains.[2] However, a more diverse fauna is known from the Cassagnau localities, in the middle part of the Marnes d’Auzas Formation.[9][3] The Cassagnau's fauna is also located near Marignac-Laspeyres and includes several fish (Lepisosteidae, Phyllodontidae, Sparidae),[9][3] Amphibians (Albanerpetontidae),[9][3] several turtles including a complete carapace of Iberoccitanemys convenarum,[9][3][10] various crocodylians including teeth of a Musturzabalsuchus-like form and a complete skull attributed to Thoracosaurus neocesariensis,[9][3][11] two squamates (an indeterminate lizard and a varanoid form[9][3] which is probably a freshwater mosasauroid[12]), a probable enantiornithe bird,[9][3] an indeterminate dromaeosaurid (known only by teeth),[9][3] an indeterminate theropod of larger size (only known by a tooth characterized by its anterior carina running on the lingual side of the teeth),[3] an indeterminate titanosaurian sauropod represented by teeth and a metacarpal[9][3] (the shape of the teeth suggests that they belong to an Atsinganosaurus-like form),[13] and an indeterminate lambeosaurine hadrosaur. The latter is exclusively represented by juvenile individuals and includes an articulated postcranial skeleton, teeth, and two left dentaries, one of them being associated with a predentary.[9][3][1] No maxilla is preserved in these specimens, so it is not possible to know if this hadrosaur belongs to Canardia or to another lambeosaurine. The Marnes d’Auzas Formation is also present in the department of Ariège, notably near the village of Mérigon. There, a coarse sandstone bed at the top of the formation, has yielded a tooth and a dentary fragment of an indeterminate hadrosaur, as well as a cervical vertebra of a giant Azhdarchid pterosaur of 9 meters wingspan.[14][3]
See also
References
- Prieto-Márquez, A.; Dalla Vecchia, F. M.; Gaete, R.; Galobart, À. (2013). Dodson, Peter (ed.). "Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis". PLOS ONE. 8 (7): e69835. Bibcode:2013PLoSO...869835P. doi:10.1371/journal.pone.0069835. PMC 3724916. PMID 23922815.
- Bilotte, M.; Laurent, Y.; Teodori, D. (2010). "Restes d'Hadrosaure dans le Crétacé terminal marin de Larcan (Petites Pyrénées, Haute-Garonne, France)". Carnets de Géologie (2010/02): 1–10. doi:10.4267/2042/32428.
- Laurent, Y. (2003). "Les faunes de vertébrés continentaux du Maastrichtien supérieur d'Europe : systématique et biodiversité". Strata (41): 1–81.
- Rozhdestvensky, A.K. (1968). "Gadrozavry Kazakhstana [Hadrosaurs of Kazakhstan]. [Upper Paleozoic and Mesozoic Amphibians and Reptiles]" (PDF). Akademia Nauk SSSR, Moscow: 97–141.
- Godefroit, P.; Alifanov, V.; Boltsky, Y. (2004). "A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan". Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre. 74Sup (74): 139–154.
- Csiki-Sava, Z.; Buffetaut, E.; Ősi, A.; Pereda-Suberbiola, X.; Brusatte, S.L. (2015). "Island life in the Cretaceous-faunal composition, biostratigraphy, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago". ZooKeys (469): 1–161. doi:10.3897/zookeys.469.8439. PMC 4296572. PMID 25610343.
- Fondevilla, V.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À.; Moncunill-Solé, B.; Köhler, M. (2018). "Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, tremp Syncline, Spain)". PLOS ONE. 13 (10): e0206287. doi:10.1371/journal.pone.0206287. PMC 6209292. PMID 30379888.
- Prieto-Márquez, A.; Fondevilla, V.; Sellés, A.G.; Wagner, J.R.; Galobart, À. (2019). "Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago". Cretaceous Research. 96: 19–37. doi:10.1016/j.cretres.2018.12.002.
- Laurent, Y.; Bilotte, M.; Le Loeuff, J. (2002). "Late Maastrichtian continental vertebrates from southwestern France: correlation with marine fauna". Palaeogeography, Palaeoclimatology, Palaeoecology. 187 (1–2): 121–135. Bibcode:2002PPP...187..121L. doi:10.1016/S0031-0182(02)00512-6.
- Pérez-García, A.; Ortega, F.; Murelaga, X. (2012). "A new genus of Bothremydidae (Chelonii, Pleurodira) in the Cretaceous of southwestern Europe". Geobios. 45 (2): 219–229. doi:10.1016/j.geobios.2011.03.001.
- Laurent, Y.; Buffetaut, E.; Le Loeuff, J. (2000). "Un crâne de Thoracosaurine (Crocodylia, Crocodylidae) dans le Maastrichtien supérieur du sud de la France". Oryctos (3): 19–27.
- Garcia, G.; Bardet, N.; Houssaye, A.; Pereda-Suberbiola, X.; Valentin, X. (2015). "Mosasauroid (Squamata) discovery in the Late Cretaceous (Early Campanian) continental deposits of Villeveyrac-L'Olivet, Southern France – Découverte de Mosasauroidea (Squamata) dans le Crétacé supérieur (Campanien inférieur) continental de Villeveyrac – L'Olivet, sud de la France". Comptes Rendus Palevol. 14 (6–7): 495–505. doi:10.1016/j.crpv.2015.05.002.
- Garcia, G.; Amicot, S.; Fournier, F.; Thouand, E.; Valentin, X. (2010). "A new Titanosaur genus (Dinosauria, Sauropoda) from the Late Cretaceous of southern France and its paleobiogeographic implications". Bulletin de la Société Géologique de France. 181 (3): 269–277. doi:10.2113/gssgfbull.181.3.269.
- Buffetaut, E.; Laurent, Y.; Le Loeuff, J.; Bilotte, M. (1997). "A terminal Cretaceous giant pterosaur from the French Pyrenees". Geological Magazine. 134 (4): 553–556. Bibcode:1997GeoM..134..553B. doi:10.1017/S0016756897007449.
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||





.jpg.webp)
