Nerve growth factor
Nerve growth factor (NGF) is a neurotrophic factor and neuropeptide primarily involved in the regulation of growth, maintenance, proliferation, and survival of certain target neurons. It is perhaps the prototypical growth factor, in that it was one of the first to be described. Since it was first isolated by Nobel Laureates Rita Levi-Montalcini and Stanley Cohen in 1956, numerous biological processes involving NGF have been identified, two of them being the survival of pancreatic beta cells and the regulation of the immune system.




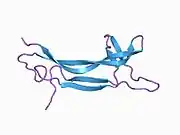
Structure
NGF is initially in a 7S, 130-kDa complex of 3 proteins - Alpha-NGF, Beta-NGF, and Gamma-NGF (2:1:2 ratio) when expressed. This form of NGF is also referred to as proNGF (NGF precursor). The gamma subunit of this complex acts as a serine protease, and cleaves the N-terminal of the beta subunit, thereby activating the protein into functional NGF.
The term nerve growth factor usually refers to the 2.5S, 26-kDa beta subunit of the protein, the only component of the 7S NGF complex that is biologically active (i.e. acting as signaling molecules).
Function
As its name suggests, NGF is involved primarily in the growth, as well as the maintenance, proliferation, and survival of nerve cells (neurons). In fact, NGF is critical for the survival and maintenance of sympathetic and sensory neurons, as they undergo apoptosis in its absence.[5] However, several recent studies suggest that NGF is also involved in pathways besides those regulating the life cycle of neurons.
Neuronal proliferation
NGF can drive the expression of genes such as bcl-2 by binding to the Tropomyosin receptor kinase A, which stimulates the proliferation and survival of the target neuron.
High affinity binding between proNGF, sortilin, and p75NTR can result in either survival or programmed cell death. Study results indicate that superior cervical ganglia neurons that express both p75NTR and TrkA die when treated with proNGF,[6] while NGF treatment of these same neurons results in survival and axonal growth. Survival and PCD mechanisms are mediated through adaptor protein binding to the death domain of the p75NTR cytoplasmic tail. Survival occurs when recruited cytoplasmic adaptor proteins facilitate signal transduction through tumor necrosis factor receptor members such as TRAF6, which results in the release of nuclear factor κB (NF-κB) transcription activator.[7] NF-κB regulates nuclear gene transcription to promote cell survival. Alternatively, programmed cell death occurs when TRAF6 and neurotrophin receptor interacting factor (NRIF) are both recruited to activate c-Jun N-terminal kinase (JNK); which phosphorylates c-Jun. The activated transcription factor c-Jun regulates nuclear transcription via AP-1 to increase pro-apoptotic gene transcription.[7]
Proliferation of pancreatic beta cells
There is evidence that pancreatic beta cells express both the TrkA and p75NTR receptors of NGF. It has been shown that the withdrawal of NGF induces apoptosis in pancreatic beta cells, signifying that NGF may play a critical role in the maintenance and survival of pancreatic beta cells.[8]
Regulation of the immune system
NGF plays a critical role in the regulation of both innate and acquired immunity. In the process of inflammation, NGF is released in high concentrations by mast cells, and induces axonal outgrowth in nearby nociceptive neurons. This leads to increased pain perception in areas under inflammation. In acquired immunity, NGF is produced by the Thymus as well as CD4+ T cell clones, inducing a cascade of maturation of T cells under infection.[9]
Ovulation
NGF is abundant in seminal plasma. Recent studies have found that it induces ovulation in some mammals e.g. “induced” ovulators, such as llamas. Surprisingly, research showed that these induced animals will also ovulate when semen from on-schedule or “spontaneous” ovulators, such as cattle is used. Its significance in humans is unknown. It was previously dubbed ovulation-inducing factor (OIF) in semen before it was identified as beta-NGF in 2012.[10]
Romantic love
Studies have found that the concentration of NGF in the blood plasma is significantly higher in individuals who have been in a romantic relationship for less than 12 months [227 (14) pg/ml], than those who are either not in a romantic relationship [149 (12) pg/ml] or have been in one for more than 12 months [123 (10) pg/ml].[11]
NGF can indirectly stimulate the expression of adrenocorticotrophic hormone (ACTH) in the hypothalamic-pituitary-adrenal axis (HPA) by increasing vasopressin secretion. ACTH binds to the MC2 receptor in the zona fasciculata of the adrenal cortex, and stimulates secretion of the stress hormone cortisol.[12] This rapid increase of cortisol in the blood plasma can induce feelings of euphoria, which may explain the initial "rush" of falling in love.[13] Studies show that ACTH can in turn stimulate NGF secretion in both the cerebral cortex and the hypothalamus.
Mechanism of action
NGF binds with at least two classes of receptors: the tropomyosin receptor kinase A (TrkA) and low-affinity NGF receptor (LNGFR/p75NTR). Both are associated with neurodegenerative disorders.
When NGF binds to the TrkA receptor, it drives the homodimerization of the receptor, which in turn causes the autophosphorylation of the tyrosine kinase segment.[14] The tropomyosin receptor kinase A receptor has five extracellular domains, and the fifth domain is sufficient in binding NGF.[15] Once bound, the complex undergoes endocytosis and activates the NGF transcriptional program, following itwo major pathways, the Ras/MAPK pathway and the PI3K/Akt pathway.[14] The binding of NGF to TrkA also leads to the activation of PI 3-kinase, ras, and PLC signaling pathways.[16] Alternatively, the p75NTR receptor can form a heterodimer with TrkA, which has higher affinity and specificity for NGF.
Studies suggest that NGF circulates throughout the entire body via the blood plasma, and is important for the overall maintenance of homeostasis.[17]
Neuron survival
Binding interaction between NGF and the TrkA receptor facilitates receptor dimerization and tyrosine residue phosphorylation of the cytoplasmic tail by adjacent Trk receptors.[18] Trk receptor phosphorylation sites operate as Shc adaptor protein docking sites, which undergo phosphorylation by the TrkA receptor[7] Once the cytoplasmic adaptor protein (Shc) is phosphorylated by the receptor cytoplasmic tail, cell survival is initiated through several intracellular pathways.
One major pathway leads to the activation of the serine/threonine kinase, Akt. This pathway begins with the Trk receptor complex-recruitment of a second adaptor protein called growth factor-receptor bound protein-2 (Grb2) along with a docking protein called Grb2-associated Binder-1 (GAB1).[7] Subsequently, phosphatidylinositol-3 kinase (PI3K) is activated, resulting in Akt kinase activation.[7] Study results have shown that blocking PI3K or Akt activity results in death of sympathetic neurons in culture, regardless of NGF presence.[19] However, if either kinase is constitutively active, neurons survive even without NGF.[19]
A second pathway contributing to cell survival occurs through activation of the mitogen-activated protein kinase (MAPK) kinase. In this pathway, recruitment of a guanine nucleotide exchange factor by the adaptor and docking proteins leads to activation of a membrane-associated G-protein known as Ras.[7] The guanine nucleotide exchange factor mediates Ras activation through the GDP-GTP exchange process. The active Ras protein phosphorylates several proteins, along with the serine/threonine kinase, Raf.[7] Raf in turn activates the MAPK cascade to facilitate ribosomal s6 kinase (RSK) activation and transcriptional regulation.[7]
Both Akt and RSK, components of the PI3K-Akt and MAPK pathways respectively, act to phosphorylate the cyclic AMP response element binding protein (CREB) transcription factor.[7] Phosphorylated CREB translocates into the nucleus and mediates increased expression of anti-apoptotic proteins,[7] thus promoting NGF-mediated cell survival. However, in the absence of NGF, the expression of pro-apoptotic proteins is increased when the activation of cell death-promoting transcription factors such as c-Jun are not suppressed by the aforementioned NGF-mediated cell survival pathways.[7]
History
Rita Levi-Montalcini and Stanley Cohen discovered NGF in the 1950s while faculty members at Washington University in St Louis. However, its discovery, along with the discovery of other neurotrophins, was not widely recognized until 1986, when it won the Nobel Prize in Physiology or Medicine.[20][21][22]
Studies in 1971 determined the primary structure of NGF. This eventually led to the discovery of the NGF gene.
NGF is abundant in seminal plasma. Recent studies have found that it induces ovulation in some mammals.[23] Nerve Growth Factors (NGF) were initially discovered due to their actions during development, but NGF are not known to be involved in the function throughout the life of the animal.[24]
Clinical significance
Nerve growth factor prevents or reduces neuronal degeneration in animal models of neurodegenerative diseases and these encouraging results in animals have led to several clinical trials in humans.[25] NGF promotes peripheral nerve regeneration in rats.[26] The expression of NGF is increased in inflammatory diseases where it suppresses inflammation.[27] NGF appears to promote myelin repair.[28] Hence NGF may be useful for the treatment of multiple sclerosis.[29] NGF could also be involved in various psychiatric disorders, such as dementia, depression, schizophrenia, autism, Rett syndrome, anorexia nervosa, and bulimia nervosa.[30]
Dysregulation of NGF signaling has also been linked to Alzheimer's disease.[31][32][33][34][35][36] Connective tissue cells genetically engineered to synthesize and secrete NGF and implanted in patients' basal forebrains reliably pumped out NGF, which enhanced the cells’ size and their ability to sprout new neural fibers. The treatment also rescued vulnerable cells, even if they already showed the trademark signs of Alzheimer's pathology. In some patients, these beneficial effects lasted almost 10 years after the treatment. Even patients who died responded positively to the therapy. Even pathological cells with protein clumps in their cell bodies and surroundings extended their fibers toward the NGF source, maintained a healthy size and activated pro-survival signals that boosted their stress resilience. Two other patients received direct injections of modified viruses containing the NGF gene directly to their basal forebrains. This allowed the gene to express longer in the brain.[37][38]
Neurotrophins, including NGF, have been shown to affect many areas of the brain, including areas that are related to Rett syndrome, bipolar disorder, and Alzheimer's disease. Stress and/or anxiety are usually a precipitating factor in these disorders and affects levels of NGF, leading to impaired cognitive functioning.
This impaired cognitive functioning can be seen in people with schizophrenia. In treating schizophrenia, NGF levels are seen to be increased when using atypical antipsychotics, but not when using typical antipsychotics. Those using atypical medications usually report improved cognitive performance compared to those using typical antipsychotics. Higher NGF levels from the atypical antipsychotic medications may underlie the reduction in negative symptoms of schizophrenia relative to typical antipsychotics.[39]
NGF has been shown to restore learning ability in rats recovering from induced alcoholism.[40]
Rett syndrome and autism often show similar signs early in life, such as slowing development and intellectual disability. One distinguishing factor is that low levels of NGF have been found in the cerebrospinal fluid of children with Rett syndrome compared to children with Autism who have relatively normal to high levels.[41] Pharmaceutical therapies with NGF-like activity can be effective in treating Rett syndrome, including better motor and cortical functioning as well as increased social communication.[42]
Impairment of neuroplasticity and altered levels of neurotrophins are involved in bipolar disorder. NGF has been found to be decreased overall in people with bipolar disorder. More specifically, while in a manic state NGF is especially low. This leads to elevated or irritable mood with increased energy and decreased need for sleep while in a manic state. This decreased NGF may serve as a biological marker when assessing a person's present bipolar disorder state.[43] When treated with lithium, their NGF concentrations increased in the frontal cortex, limbic forebrain, hippocampus, and amygdala.[44]
An increase in cortical and subcortical NGF has been found in patients with Alzheimer's disease. Alzheimer's is a neurodegenerative disease with which dysregulation of NGF signaling has also been linked, causing impaired retrograde transport of NGF to certain areas of the brain. This impairment may be caused by an atypical production or use of receptors in the brain.[45] Stimulating NGF receptors via NGF infusion has been shown to increase blood flow and verbal episodic memory. These improvements have been longer lasting than other treatments for Alzheimer's.[42]
Also, NGF has been shown to play a role in a number cardiovascular diseases, such as coronary atherosclerosis, obesity, type 2 diabetes, and metabolic syndrome.[46] Reduced plasma levels of NGF and BDNF have been associated with acute coronary syndromes and metabolic syndromes.[47][48] NGF is known to have insulinotropic, angiogenic, and antioxidant properties. NGF suppresses food intake.
NGF has also been shown to accelerate wound healing. There is evidence that it could be useful in the treatment of skin ulcers and cornea ulcers.[49]
A mutation in the beta gene of NGF has been seen to lead to a loss of pain perception; additionally, this loss of pain is not linked with a change to CNS development or mental abilities in patients.[50] Thus, this study highlights that there may be different pathways by which the NGF gene regulates pain perception compared with other nervous system development.[50]
In some gynecological diseases, an elevated prostaglandin E2 is thought to stimulate production of NGF which contributes to the perception of pain and increased inflammation in endometriosis.[51]
Monoclonal antibodies against NGF have been used in clinical trials to modulate pain. One of these is tanezumab, another is fulranumab.
Miscellaneous
Nerve growth factor may contribute to increased longevity and mental capacity.[52] Centenarian Rita Levi-Montalcini took a daily solution in the form of eye drops, and has stated that her brain is more active now than it was four decades ago.[52] In 2014, researchers at the Medical University of South Carolina showed that NGF level is elevated in people who performed a single 20-minute yoga session involving om-chanting and thirumoolar pranayama, when compared to a control group.[53]
Interactions
Nerve growth factor has been shown to interact with [Tropomyosin receptor kinase A]][6][54][15] and p75NTR (LNGFR).[6][54]
It has recently been suggested that NGF expression may be stimulated by dehydroepiandrosterone (DHEA).[55] DHEA may also act as an agonist of both TrkA and p75NTR and activate the pathways of NGF, demonstrating neurotrophic activities similar to that of NGF.[56]
Adrenocorticotrophic hormone (ACTH) can also upregulate NGF expression in the brain.[57]
See also
- Protein targeting
- Nervous System
- VGF Nerve Growth Factor-inducible, a protein whose expression is induced by NGF
- Neurotrophin
- Neurotrophin-3
- Neurotrophin-4
- Nerve growth factor receptor
- Growth factor
- Brain-derived neurotrophic factor
- Hericium erinaceus an edible mushroom that has been shown to boost NGF
- Huperzine A an herb-derived alkaloid that seems to boost NGF
- Polygala tenuifolia a Chinese herb shown to increase NGF secretion in astrocytes
- Therapygenetics - showing how NGF genes predict treatment outcome to cognitive behavioural therapy
References
- GRCh38: Ensembl release 89: ENSG00000134259 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000027859 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Freeman RS, Burch RL, Crowder RJ, Lomb DJ, Schoell MC, Straub JA, Xie L (2004). "NGF deprivation-induced gene expression: after ten years, where do we stand?". NGF and Related Molecules in Health and Disease. Progress in Brain Research. 146. pp. 111–26. doi:10.1016/S0079-6123(03)46008-1. ISBN 978-0-444-51472-1. PMID 14699960.
- Lee R, Kermani P, Teng KK, Hempstead BL (Nov 2001). "Regulation of cell survival by secreted proneurotrophins". Science. 294 (5548): 1945–8. Bibcode:2001Sci...294.1945L. doi:10.1126/science.1065057. PMID 11729324. S2CID 872149.
- Sanes DH, Thomas AR, Harris WA (2011). "Naturally-occurring neuron death". Development of the Nervous System, Third Edition. Boston: Academic Press. pp. 171–208. ISBN 978-0-12-374539-2.
- Pierucci D, Cicconi S, Bonini P, Ferrelli F, Pastore D, Matteucci C, Marselli L, Marchetti P, Ris F, Halban P, Oberholzer J, Federici M, Cozzolino F, Lauro R, Borboni P, Marlier LN (Oct 2001). "NGF-withdrawal induces apoptosis in pancreatic beta cells in vitro". Diabetologia. 44 (10): 1281–95. doi:10.1007/s001250100650. PMID 11692177.
- Lambiase A, Bracci-Laudiero L, Bonini S, Bonini S, Starace G, D'Elios MM, De Carli M, Aloe L (Sep 1997). "Human CD4+ T cell clones produce and release nerve growth factor and express high-affinity nerve growth factor receptors". The Journal of Allergy and Clinical Immunology. 100 (3): 408–14. doi:10.1016/s0091-6749(97)70256-2. PMID 9314355.
- Ratto MH, Leduc YA, Valderrama XP, van Straaten KE, Delbaere LT, Pierson RA, Adams GP (Sep 2012). "The nerve of ovulation-inducing factor in semen". Proceedings of the National Academy of Sciences of the United States of America. 109 (37): 15042–7. Bibcode:2012PNAS..10915042R. doi:10.1073/pnas.1206273109. PMC 3443178. PMID 22908303. Lay summary – sciencenews.org.
- Emanuele, Enzo; Politi, Pierluigi; Bianchi, Marika; Minoretti, Piercarlo; Bertona, Marco; Geroldi, Diego (2006-04-01). "Raised plasma nerve growth factor levels associated with early-stage romantic love". Psychoneuroendocrinology. 31 (3): 288–294. doi:10.1016/j.psyneuen.2005.09.002. ISSN 0306-4530. PMID 16289361. S2CID 18497668.
- Otten, U.; Goedert, M.; Baumann, J. B.; Girard, J. (1981-07-27). "Stimulation of the pituitary-adrenocortical axis and induction of tyrosine hydroxylase by nerve growth factor are not dependent on mouse submaxillary gland isorenin". Brain Research. 217 (1): 207–211. doi:10.1016/0006-8993(81)90202-x. ISSN 0006-8993. PMID 6114784. S2CID 13530587.
- Valiengo, Leandro L.; Soeiro-de-Souza, Márcio G.; Marques, Andrea H.; Moreno, Doris H.; Juruena, Mário F.; Andreazza, Ana Cristina; Gattaz, Wagner F.; Machado-Vieira, Rodrigo (2012-04-01). "Plasma cortisol in first episode drug-naïve mania: Differential levels in euphoric versus irritable mood". Journal of Affective Disorders. 138 (1–2): 149–152. doi:10.1016/j.jad.2011.11.046. ISSN 0165-0327. PMC 4479259. PMID 22305430.
- Stoleru B, Popescu A, Tache D, Neamtu O, Emami G, Tataranu L, Buteica A, Dricu A, Purcaru S (2013). "Tropomyosin-Receptor-Kinases Signaling in the Nervous System". Maedica. 8 (1): 43–48. PMC 3749761. PMID 24023598.
- Wiesmann C, Ultsch MH, Bass SH, de Vos AM (September 1999). "Crystal structure of nerve growth factor in complex with the ligand-binding domain of the TrkA receptor". Nature. 401 (6749): 184–8. Bibcode:1999Natur.401..184W. doi:10.1038/43705. PMID 10490030. S2CID 4337786.
- Marlin MC, Li G (2015). "Biogenesis and function of the NGF/TrkA signaling endosome". International Review of Cell and Molecular Biology. 314: 239–57. doi:10.1016/bs.ircmb.2014.10.002. ISBN 9780128022832. PMC 4307610. PMID 25619719.
- Levi-Montalcini R (2004). "The nerve growth factor and the neuroscience chess board". Progress in Brain Research. 146: 525–7. doi:10.1016/s0079-6123(03)46033-0. ISBN 9780444514721. PMID 14699984.
- Kaplan DR, Martin-Zanca D, Parada LF (Mar 1991). "Tyrosine phosphorylation and tyrosine kinase activity of the trk proto-oncogene product induced by NGF". Nature. 350 (6314): 158–60. Bibcode:1991Natur.350..158K. doi:10.1038/350158a0. PMID 1706478. S2CID 4241996.
- Crowder RJ, Freeman RS (Apr 1998). "Phosphatidylinositol 3-kinase and Akt protein kinase are necessary and sufficient for the survival of nerve growth factor-dependent sympathetic neurons". The Journal of Neuroscience. 18 (8): 2933–43. doi:10.1523/JNEUROSCI.18-08-02933.1998. PMC 6792598. PMID 9526010.
- The 1986 Nobel Prize in Physiology or Medicine for discoveries of growth factors
- Presentation Speech by Professor Kerstin Hall The Nobel Prize in Physiology or Medicine 1986
- Rita Levi-Montalcini – Nobel Lecture
- Ovulation spurred by newfound semen ingredient
- Adelman, George. Encyclopedia of Neuroscience . Boston: Birkhhaeuser, 1987. Print.
- Tuszynski MH, Blesch A (2004). "Nerve growth factor: from animal models of cholinergic neuronal degeneration to gene therapy in Alzheimer's disease". Progress in Brain Research. 146: 441–9. doi:10.1016/s0079-6123(03)46028-7. ISBN 9780444514721. PMID 14699979.
- Sun W, Sun C, Lin H, Zhao H, Wang J, Ma H, Chen B, Xiao Z, Dai J (Sep 2009). "The effect of collagen-binding NGF-beta on the promotion of sciatic nerve regeneration in a rat sciatic nerve crush injury model". Biomaterials. 30 (27): 4649–56. doi:10.1016/j.biomaterials.2009.05.037. PMID 19573907.
- Freund V, Frossard N (2004). "Expression of nerve growth factor in the airways and its possible role in asthma". Progress in Brain Research. 146: 335–46. doi:10.1016/S0079-6123(03)46021-4. ISBN 9780444514721. PMID 14712791.
- Althaus HH (2004). "Remyelination in multiple sclerosis: a new role for neurotrophins?". Progress in Brain Research. 146: 415–32. doi:10.1016/S0079-6123(03)46026-3. ISBN 9780444514721. PMID 14699977.
- Villoslada P, Genain CP (2004). "Role of nerve growth factor and other trophic factors in brain inflammation". Progress in Brain Research. 146: 403–14. doi:10.1016/S0079-6123(03)46025-1. ISBN 9780444514721. PMID 14699976.
- Chaldakov GN, Tonchev AB, Aloe L (2009). "NGF and BDNF: from nerves to adipose tissue, from neurokines to metabokines". Rivista di Psichiatria. 44 (2): 79–87. PMID 20066808.
- Counts SE, Mufson EJ (Apr 2005). "The role of nerve growth factor receptors in cholinergic basal forebrain degeneration in prodromal Alzheimer disease". Journal of Neuropathology and Experimental Neurology. 64 (4): 263–72. doi:10.1093/jnen/64.4.263. PMID 15835262.
- Hempstead BL (Feb 2006). "Dissecting the diverse actions of pro- and mature neurotrophins". Current Alzheimer Research. 3 (1): 19–24. doi:10.2174/156720506775697061. PMID 16472198.
- Allen SJ, Dawbarn D (Feb 2006). "Clinical relevance of the neurotrophins and their receptors". Clinical Science. 110 (2): 175–91. doi:10.1042/CS20050161. PMID 16411894.
- Barry SR (Dec 1991). "Clinical implications of basic neuroscience research. II: NMDA receptors and neurotrophic factors". Archives of Physical Medicine and Rehabilitation. 72 (13): 1095–101. PMID 1660257.
- Hefti F, Schneider LS (1991). "Nerve growth factor and Alzheimer's disease". Clinical Neuropharmacology. 14 Suppl 1: S62-76. doi:10.1097/00002826-199114001-00008. PMID 1913710. S2CID 45315301.
- Olson L (Nov 1993). "NGF and the treatment of Alzheimer's disease". Experimental Neurology. 124 (1): 5–15. doi:10.1006/exnr.1993.1167. PMID 8282080. S2CID 43552000.
- Fanon, Shelly (September 6, 2015). "Gene Therapy May Be Our Newest — and Most Effective — Weapon Against Alzheimer's Disease". Singularity HUB. Retrieved 2015-09-10.
- Tuszynski MH, Yang JH, Barba D, U HS, Bakay RA, Pay MM, Masliah E, Conner JM, Kobalka P, Roy S, Nagahara AH (Aug 2015). "Nerve Growth Factor Gene Therapy: Activation of Neuronal Responses in Alzheimer Disease". JAMA Neurology. 72 (10): 1139–47. doi:10.1001/jamaneurol.2015.1807. PMC 4944824. PMID 26302439.
- Parikh V, Evans DR, Khan MM, Mahadik SP (Apr 2003). "Nerve growth factor in never-medicated first-episode psychotic and medicated chronic schizophrenic patients: possible implications for treatment outcome". Schizophrenia Research. 60 (2–3): 117–23. doi:10.1016/S0920-9964(02)00434-6. PMID 12591576. S2CID 32114191.
- Lukoyanov NV, Pereira PA, Paula-Barbosa MM, Cadete-Leite A (Jan 2003). "Nerve growth factor improves spatial learning and restores hippocampal cholinergic fibers in rats withdrawn from chronic treatment with ethanol". Experimental Brain Research. 148 (1): 88–94. doi:10.1007/s00221-002-1290-7. PMID 12478399. S2CID 19599008.
- Riikonen R, Vanhala R (Mar 1999). "Levels of cerebrospinal fluid nerve-growth factor differ in infantile autism and Rett syndrome". Developmental Medicine and Child Neurology. 41 (3): 148–52. doi:10.1111/j.1469-8749.1999.tb00573.x. PMID 10210246. S2CID 221649358.
- Gorbachevskaya N, Bashina V, Gratchev V, Iznak A (Dec 2001). "Cerebrolysin therapy in Rett syndrome: clinical and EEG mapping study". Brain & Development. 23 Suppl 1: S90-3. doi:10.1016/S0387-7604(01)00349-7. PMID 11738849. S2CID 23240417.
- Barbosa IG, Huguet RB, Neves FS, Reis HJ, Bauer ME, Janka Z, Palotás A, Teixeira AL (Apr 2011). "Impaired nerve growth factor homeostasis in patients with bipolar disorder". The World Journal of Biological Psychiatry. 12 (3): 228–32. doi:10.3109/15622975.2010.518629. PMID 20923384. S2CID 12821162.
- Machado-Vieira R, Manji HK, Zarate CA (Jun 2009). "The role of lithium in the treatment of bipolar disorder: convergent evidence for neurotrophic effects as a unifying hypothesis". Bipolar Disorders. 11 Suppl 2: 92–109. doi:10.1111/j.1399-5618.2009.00714.x. PMC 2800957. PMID 19538689.
- Scott SA, Mufson EJ, Weingartner JA, Skau KA, Crutcher KA (Sep 1995). "Nerve growth factor in Alzheimer's disease: increased levels throughout the brain coupled with declines in nucleus basalis". The Journal of Neuroscience. 15 (9): 6213–21. doi:10.1523/JNEUROSCI.15-09-06213.1995. PMC 6577665. PMID 7666203.
- Chaldakov GN, Fiore M, Stankulov IS, Manni L, Hristova MG, Antonelli A, Ghenev PI, Aloe L (2004). "Neurotrophin presence in human coronary atherosclerosis and metabolic syndrome: a role for NGF and BDNF in cardiovascular disease?". Progress in Brain Research. 146: 279–89. doi:10.1016/S0079-6123(03)46018-4. ISBN 9780444514721. PMID 14699970.
- Chaldakov GN, Fiore M, Tonchev AB, Dimitrov D, Pancheva R, Rancic G, Aloe L (2007). "Homo obesus: a metabotrophin-deficient species. Pharmacology and nutrition insight". Current Pharmaceutical Design. 13 (21): 2176–9. doi:10.2174/138161207781039616. PMID 17627549.
- Manni L, Nikolova V, Vyagova D, Chaldakov GN, Aloe L (Jun 2005). "Reduced plasma levels of NGF and BDNF in patients with acute coronary syndromes". International Journal of Cardiology. 102 (1): 169–71. doi:10.1016/j.ijcard.2004.10.041. PMID 15939120.
- Kawamoto K, Matsuda H (2004). "Nerve growth factor and wound healing". Progress in Brain Research. 146: 369–84. doi:10.1016/S0079-6123(03)46023-8. ISBN 9780444514721. PMID 14699974.
- Einarsdottir E, Carlsson A, Minde J, Toolanen G, Svensson O, Solders G, Holmgren G, Holmberg D, Holmberg M (2004). "A mutation in the nerve growth factor beta gene (NGFB) causes loss of pain perception". Human Molecular Genetics. 13 (8): 799–805. doi:10.1093/hmg/ddh096. PMID 14976160.
- Giudice LC (Jun 2010). "Clinical practice. Endometriosis". The New England Journal of Medicine. 362 (25): 2389–98. doi:10.1056/NEJMcp1000274. PMC 3108065. PMID 20573927.
- Popham P (2009-04-25). "Is this the secret of eternal life? - Science - News - The Independent". The Independent. Retrieved 2012-03-02.
- Balasubramanian S, Mintzer JE, Wahlquist AE (Jan 2015). "Induction of salivary nerve growth factor by Yogic breathing: a randomized controlled trial". International Psychogeriatrics / IPA. 27 (1): 168–70. doi:10.1017/S1041610214001616. PMC 4501575. PMID 25101659.
- Nykjaer A, Lee R, Teng KK, Jansen P, Madsen P, Nielsen MS, Jacobsen C, Kliemannel M, Schwarz E, Willnow TE, Hempstead BL, Petersen CM (Feb 2004). "Sortilin is essential for proNGF-induced neuronal cell death". Nature. 427 (6977): 843–8. Bibcode:2004Natur.427..843N. doi:10.1038/nature02319. PMID 14985763. S2CID 4343450.
- Rahmani A, Shoae-Hassani A, Keyhanvar P, Kheradmand D, Darbandi-Azar A (2011). "Dehydroepiandrosterone stimulates nerve growth factor and brain derived neurotrophic factor in cortical neurons". Advances in Pharmacological Sciences. 2013: 506191. doi:10.1155/2013/506191. PMC 3867952. PMID 24381588.
- Lazaridis I, Charalampopoulos I, Alexaki VI, Avlonitis N, Pediaditakis I, Efstathopoulos P, Calogeropoulou T, Castanas E, Gravanis A (Apr 2011). "Neurosteroid dehydroepiandrosterone interacts with nerve growth factor (NGF) receptors, preventing neuronal apoptosis". PLOS Biology. 9 (4): e1001051. doi:10.1371/journal.pbio.1001051. PMC 3082517. PMID 21541365.
- Mocchetti, Spiga, Hayes, Isackson, Colangelo, Italo, Giulio, Valerie, Paul, Annamaria (March 15, 1996). "Glucocorticoids Differently Increase Nerve Growth Factor and Basic Fibroblast Growth Factor in the Rat Brain" (PDF). The Journal of Neuroscience. doi:10.1523/JNEUROSCI.16-06-02141.1996. S2CID 11078370. Retrieved September 9, 2015.CS1 maint: multiple names: authors list (link)
External links
- Nerve+Growth+Factor at the US National Library of Medicine Medical Subject Headings (MeSH)
- NCBI: nerve growth factor (beta polypeptide)
- NGF for corneal therapeutic purposes
- Overview of all the structural information available in the PDB for UniProt: P01138 (Human Beta-nerve growth factor) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: P01139 (Mouse Beta-nerve growth factor) at the PDBe-KB.
PDB gallery | |
|---|---|
|