Timeline of dromaeosaurid research
This timeline of dromaeosaurid research is a chronological listing of events in the history of paleontology focused on the dromaeosaurids, a group of sickle-clawed, bird-like theropod dinosaurs including animals like Velociraptor. Since the Native Americans of Montana used the sediments of the Cloverly Formation to produce pigments, they may have encountered remains of the dromaeosaurid Deinonychus hundreds of years before these fossils came to the attention of formally trained scientists.[1]
.jpg.webp)
In 1922 Matthew and Brown named the new genus and species Dromaeosaurus albertensis, considering it a new type within the family Deinodontidae, a now-defunct family name that once applied to the tyrannosaurs. Not long after, Velociraptor was discovered in Mongolia by the Central Asiatic Expedition. Dromaeosaur research was fairly quiet until the 1960s, when John Ostrom described the new genus and species Deinonychus antirrhopus.[2] This discovery played a major role in setting off the Dinosaur Renaissance because Deinonychus was obviously a vigorous, active animal, and exhibited characteristics linking it to the origin of birds. As such it brought support for controversial reinterpretations of dinosaurs as warm-blooded and ancestral to birds.[3] Its distinct nature and similarity to Dromaeosaurus led Ostrom to follow Edwin Colbert and Dale Russel's suggestion that the Dromaeosaurinae be regarded as its own family separate from the Deinodontidae.[4]
After Ostrom's initial research on Deinonychus, evidence continued to mount for a close evolutionary relationship between dromaeosaurids and birds.[5] The dromaeosaurid Sinornithosaurus milennii, described in 1999 by Xu, Wang, and Wu, is a notable example as the fine-grained Chinese limestone from which it was collected preserved its life covering of feathers.[2] Discoveries of feathered dromaeosaurids continued into the 2000s. Xu, Zhou, and Wang named the new genus Microraptor in 2000.[6] Three years later, Xu and others would report a new species in this genus that exhibited a bizarre "four winged" body plan with long pennaceous flight feathers on both its front and hind limbs.[7]
Prescientific

- The Crow people and other Native American groups inhabiting Montana used to use rocks from the Cloverly Formation to make red pigments. Since the red pigments are richest in the same layers of the formation that preserve dinosaur fossils, it is likely that Native Americans encountered Deinonychus fossils long before scientifically trained paleontologists.[1]
19th century
20th century

1910s
- Barnum Brown collected a partial skull and foot in Late Cretaceous rocks that would later serve as the type specimen of Dromaeosaurus albertensis.[2]
1920s

- Matthew and Brown described the new genus and species Dromaeosaurus albertensis and named the Dromaeosauridae.[6]
- Henry Fairfield Osborn described the new genus and species Velociraptor mongoliensis.[8]
1960s

- John Ostrom described the new genus and species Deinonychus antirrhopus.[6] This description provided the scientific literature with its first detailed information about the anatomy of the dromaeosaurid body, as previous work focused on the skull.[2] He proposed that Deinonychus killed its prey with the enlarged, sharply curved claws on the second toe of its feet.[9]
- Ostrom further described the anatomy of Deinonychus antirrhopus, providing further details about the anatomy of its body.[2] Ostrom proposed that Deinonychus only walked on two of its toes.[9]
1970s
.jpg.webp)
- Zofia Kielan-Jaworowska and Rinchen Barsbold reported the associated remains of a Velociraptor and Protoceratops apparently killed and preserved while fighting.[9]
- Ostrom first discussed the apparent close evolutionary relationship between Deinonychus antirrhopus and birds.[5]
- Ostrom published further arguments building a case for a close evolutionary relationship between dromaeosaurids and birds.[5]

- Ostrom published further arguments building a case for a close evolutionary relationship between dromaeosaurids and birds.[5]
- Sues described the new genus and species Saurornitholestes langstoni.[6]
- Powell described the new genus and species Unquillosaurus ceibalii.[8]
1980s

- Jensen described the new genus and species Palaeopteryx thomsoni.[8]
- Osmolska described the new genus and species Hulsanpes perlei.[8]

- Rinchen Barsbold divided the Dromaeosauridae into two subfamilies; the Dromaeosaurinae and Velociraptorinae.[5]
- Barsbold described the new genus and species Adasaurus mongoliensis.[6] Unlike most dromaeosaurs, famous for the "killing claws" on their second toe, the second toe claw of A. mongoliensis was relatively small and not sharply curved. It is similar to those of troodontids.[9]
- Ostrom published further arguments building a case for a close evolutionary relationship between dromaeosaurids and birds.[5]
1990s
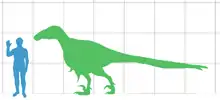
- Ostrom published further arguments building a case for a close evolutionary relationship between dromaeosaurids and birds.[5]
- Kirkland, Burge, and Gaston described the new genus and species Utahraptor ostrommaysi.[6]
- Osmolska challenged Kielan-Jaworowska and Barsbold's 1972 interpretation of an associated skeleton of Velociraptor and Protoceratops as being of two animals killed and preserved while locked in mortal combat. She reinterpreted the association as a Velociraptor that had been killed while scavenging an already dead Protoceratops.[9]

- Ostrom speculated that Deinonychus hunted in packs to subdue prey larger than itself based on an association between several Deinonychus skeletons and a specimen of the ornithopod Tenontosaurus.[9]
- Currie followed Barsbold's division of Dromaeosauridae into Dromaeosaurinae and Velociraptorinae. However, he proposed differing definitions for these subfamilies and argued that they had different members than were included under Barsbold's classification scheme.[5]
- Maxwell and Ostrom reported on the association of Deinonychus teeth and Tenontosaurus remains and argued that these provided further support for the hypothesis that Deinonychus was a pack hunter.[9]
- Unwin and others followed the fighting-to-the-death interpretation of the fighting dinosaurs specimen.[9]

- Novas and Puerta described the new genus and species Unenlagia comahuensis.[6]
- Chatterjee argued that dromaeosaurids lived in trees based on the anatomy of their hands and the backward orientation of the pubic bones in their pelvis.[9]
- Reid examined histological sections of the bones of Saurornitholestes to see whether or not they were consistent with the idea that dromaeosaurids were warm blooded. However, the results of his study were inconclusive.[9]
- Ruben and others attempted to reconstruct the way air would flow through the snouts of dromaeosaurids during respiration to see whether or not it was consistent with the airflow patterns of modern cold or warm-blooded animals. They found that the probable course of airflow through the snout of a living, breathing dromaeosaurid would have been more consistent with that of a cold blooded animal. However, the anatomical reconstructions the researchers used to draw these conclusions have been criticized for positioning the choana too far forward and for excluding the animals' secondary palate.[9]
- Le Leouff and Buffetaut described the new genus and species Variraptor mechinorum.[8]
- Novas described the new genus and species Megaraptor namunhuaiquii.[8]

- D. L. Brinkman and others reinterpreted the association between several Deinonychus specimens and a Tenontosaurus as preserving the aftermath of an ancient feeding frenzy rather than as evidence of pack hunting behavior in Deinonychus as argued by Ostrom.[9]
- Clark and others argued that it was impossible to know whether or not Velociraptor was a predator of Protoceratops based on the famous "fighting dinosaurs" specimen, since this was only one specimen.[9]
- Altangerel Perle, Norell and Clark described the new genus and species Achillobator giganticus.[6]
- Xu, Wang, and Wu described the new genus and species Sinornithosaurus millenii.[6] The specimen actually preserved the remains of the feathers that covered the animal in life. The presence of a feathery covering is consistent with the idea that dromaeosaurids were warm-blooded.[9] They also performed a cladistic analysis of the Dromaeosauridae, finding more support for dividing the family into Dromaeosaurinae and Velociraptorinae. The researchers found both groups to be monophyletic clades, with Sinornithosaurus milenii the sister group to both, and therefore the most primitive known dromaeosaurid.[10]
21st century
2000s
- Burnham and others described the new genus and species Bambiraptor feinbergorum.[6]
- Xu, Zhou and Wang described the new genus and species Microraptor zhaoianus.[6]
- Allain and Taquet described the new genus and species Pyroraptor olympius.[8]
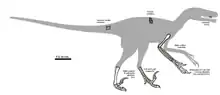


- A. R. Jacobsen published a description of a dentary referred to Saurornitholestes with tooth marks.[11] The specimen was preserved in the Dinosaur Park Formation.[12] Although a specific identification cannot be made, the shape of the preserved serration marks implicate a juvenile individual of one of the formation's tyrannosaurids, like Gorgosaurus, Daspletosaurus, or Aublysodon.[13] All of the marks on the jawbone seem to have been left by the same animal because the serration marks all share the same morphology.[14]
- Kevin Padian, Ji Qiang and Ji Shu-an published a review of known feathered dinosaurs and their implications for the origin of flight.[15] The authors observed that many aspects of the distribution of feather homologues in the dinosaur family tree met the expectations of earlier phylogenetic hypotheses, including a gradual transition from primitive filaments in Sinosauropteryx to the shared filaments and "rudimentary" true feathers in Caudipteryx and Protarchaeopteryx, to flight feathers in Archaeopteryx.[16] However, they also noted, only filamentous protofeathers had been observed in dromaeosaurs like Sinornithosaurus, when Caudipteryx-style true feathers would be expected due to the family's phylogenetic position.[17] Also noted were the lack of known integumentary structures among troodontids.[17] The authors proposed that either science's understanding of coelurosaur evolution is faulty or feathers evolved multiple times.[17]
Cladogram of feathered dinosaurs from Padian et al. 2001
Feathered Dinosaurs
Coelurosaurs
Maniraptora
Aves
Neornithes (P)
Enantiornithes (P)
Confuciusornis (P)
Archaeopteryx (P)
Dromaeosaurs (F)
Caudipteryx (F)
Therizinosaurs (F)
Sinosauropteryx (F)
Taxa marked with a P were known to bear plumulaceous or pennaceous feathers at the time of the study. Taxa marked with F were known to bear simple filamentous integumentary structures.
- Hwang and others performed a cladistic analysis of the Dromaeosauridae.[10]
- Xu and others performed a cladistic analysis of the Dromaeosauridae.[10]

- Czerkas and others described the new genus and species Cryptovolans pauli.[18]
- Christiansen and Bonde described the new genus and species Dromaeosauroides bornholmensis.[19]
- Currie and Varricchio described the new genus and species Atrociraptor marshalli.[20]

- Xu and Wang described the new genus and species Graciliraptor lujiatunensis.[21]
- Makovicky, Apesteguía and Agnolin described the new genus and species Buitreraptor gonzalezorum.[22]
- Novas and Pol described the new genus and species Neuquenraptor argentinus.[23]
- Norell and others described the new genus and species Tsaagan mangas.[24]
- Lü and others described the new genus and species Luanchuanraptor henanensis.[25]

- Turner and others described the new genus and species Mahakala omnogovae.[26]
- A. S. Turner, S. H. Hwang, and M. A. Norell described the new genus and species Shanag ashile.[27]
- Novas et al. described the new genus and species Austroraptor cabazai.[28]
- Longrich and Currie described the new genus and species Hesperonychus elizabethae.[29]
2010s
- Xu and others described the new genus and species Linheraptor exquisitus.[30]
- Zheng and others described the new genus and species Tianyuraptor ostromi.[31]
- Porfiri, Calvo and Santos described the new genus and species Pamparaptor micros.[32]
- Gong and others described the new species Microraptor hanqingi.[33]

- Senter and others described the new genus and species Yurgovuchia doellingi.[34]
- D. C. Evans, D. W. Larson, and P. J. Currie described the new genus and species Acheroraptor temertyorum.[35]
- Han and others described the new genus and species Changyuraptor yangi.[36]
- Jasinski described the new species Saurornitholestes sullivani.[37]
- Lü and Brusatte described the new genus and species Zhenyuanlong suni.[38]
- DePalma et al. described the new genus and species Dakotaraptor steini.[39]
Cladogram of dromaeosaurids from Lü and Brusatte 2015
Dromaeosauridae
Zhenyuanlong
Eudromaeosauria
Dromaeosaurinae
- Bell and Currie described the new genus and species Boreonykus certekorum.[40]
- Xu and others described the new genus and species Zhongjianosaurus yangi.
- Cau and others described the new genus and species Halszkaraptor escuilliei.
- An ungual phalanx of a dromaeosaurid theropod is described from the Blagoveshchensk area (Russia) by Bolotskii, Bolotskii & Sorokin (2019).[41]
Footnotes
- Mayor (2005); "Crow Fossil Collections," pages 272–273.
- Norell and Makovicky (2004); "Introduction", page 196.
- Horner (2001); "History of Dinosaur Collecting in Montana," pages 53–54.
- Ostrom (1969); "6. Affinities of Deinonychus," pages 147–148.
- Norell and Makovicky (2004); "Systematics and Evolution", page 206.
- Norell and Makovicky (2004); "Table 10.1: Dromaeosauridae", page 198.
- Xu et al. (2003); in passim pages 335–340.
- Norell and Makovicky (2004); "Table 10.1: Dromaeosauridae", page 199.
- Norell and Makovicky (2004); "Paleoecology", page 209.
- Norell and Makovicky (2004); "Systematics and Evolution", page 207.
- Jacobsen (2001); "Abstract," page 58.
- Jacobsen (2001); "Introduction," page 59.
- Jacobsen (2001); "Discussion," page 61.
- Jacobsen (2001); "Discussion," page 60.
- Padian, Ji, and Ji (2001); "Abstract," page 117.
- Padian, Ji, and Ji (2001); "Conclusions," pages 131–132.
- Padian, Ji, and Ji (2001); "Conclusions," page 132.
- Czerkas et al. (2002); "Abstract," page 96.
- Christiansen and Bonde (2003); "Abstract," page 287.
- Currie and Varricchio (2004); "Abstract," page 112.
- Xu and Wang (2004); "Abstract," page 11.
- Makovicky, Apesteguia and Agnolin (2005); "Abstract," page 1007.
- Novas and Pol (2005); "Abstract," page 858.
- Norell et al. (2006); "Abstract," page 1.
- Lü et al. (2007); "Abstract," page 777.
- Turner et al. (2007); "Abstract," page 1378.
- Turner, Hwang, and Norell (2007); "Abstract," page 1.
- Novas et al. (2008); "Abstract," page 1101.
- Longrich and Currie (2009); "Abstract," page 5002.
- Xu et al. (2010); "Abstract," page 1.
- Zheng et al. (2010); "Abstract," page 211.
- Porfiri, Calvo and Santos (2011); "Abstract," page 109.
- Gong et al. (2012); "Abstract," page 1.
- Senter et al. (2012); "Abstract," page 58.
- Evans, Larson, and Currie (2013); "Abstract," page 1041.
- Han et al. (2014); "Abstract".
- Jasinski (2015); "Abstract", page 79.
- Lü and Brusatte (2015); "Abstract", page 1.
- DePalma et al. (2015); in passim.
- Bell and Currie (2015); in passim.
- I. Yu. Bolotskii; Yu. L. Bolotskii; A. P. Sorokin (2019). "The first find of an ungual phalanx of a dromaeosaurid dinosaur (Dinosauria: Dromaeosauridae) from the Blagoveshchensk area of Late Cretaceous dinosaurs (Amur Region, Russia)". Doklady Earth Sciences. 484 (1): 18–20. Bibcode:2019DokES.484...18B. doi:10.1134/S1028334X19010100. S2CID 134803475.
References
- Phil R. Bell & Philip J. Currie (2015). "A high-latitude dromaeosaurid, Boreonykus certekorum, gen. et sp. nov. (Theropoda), from the upper Campanian Wapiti Formation, west-central Alberta". Journal of Vertebrate Paleontology. Online edition: e1034359. doi:10.1080/02724634.2015.1034359. S2CID 131680329.
- Christiansen P. & Bonde N. (2003). "The first dinosaur from Denmark". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 227 (2): 287–299.
- Z., Csiki; Vremir, M.; Brusatte, S. L.; Norell, M. A. (2010). "An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania". Proceedings of the National Academy of Sciences of the United States of America. 107 (35): 15357–15361. Bibcode:2010PNAS..10715357C. doi:10.1073/pnas.1006970107. PMC 2932599. PMID 20805514.
- Currie, P. J.; Varricchio, D. J. (2004). "A new dromaeosaurid from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta, Canada" (PDF). In Currie, P. J.; Koppelhus, E. B.; Shugar, M. A.; Wright, J. L. (eds.). Feathered Dragons. Life of the Past. Indianapolis: Indiana University Press. pp. 112–132.
- Czerkas, S. A.; Zhang, D.; Li, J; Li, Y (2002). "Flying dromaeosaurs". In Czerkas, S. J. (ed.). Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal. 1. Blanding: The Dinosaur Museum. pp. 96–126.
- DePalma, Robert A.; Burnham, David A.; Martin, Larry D.; Larson, Peter L.; Bakker, Robert T. (2015). "The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontological Contributions. 14: 1–16.
- Evans, DC; Larson, DW; Currie, PJ (2013). "A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America". Naturwissenschaften. 100 (11): 1041–1049. Bibcode:2013NW....100.1041E. doi:10.1007/s00114-013-1107-5. PMID 24248432. S2CID 14978813.
- Gang Han, Luis M. Chiappe, Shu-An Ji, Michael Habib, Alan H. Turner, Anusuya Chinsamy, Xueling Liu & Lizhuo Han (15 July 2014). "A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance". Nature Communications. 5: 4382. Bibcode:2014NatCo...5.4382H. doi:10.1038/ncomms5382. PMID 25025742.CS1 maint: uses authors parameter (link)
- Gong; Martin; Burnham; Falk; Hou (2012). "A new species of Microraptor from the Jehol Biota of northeastern China". Palaeoworld. 21 (2): 81–91. doi:10.1016/j.palwor.2012.05.003.
- Horner, John R. (2001). Dinosaurs Under the Big Sky. Mountain Press Publishing Company. ISBN 0-87842-445-8.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A. (2004). "Hadrosauridae". In Weishampel, D. B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (2 ed.). Berkeley: University of California Press. pp. 438–463. ISBN 978-0520254084.
- Jacobsen, A. R. (2001). "Tooth-marked small theropod bone: An extremely rare trace". In Tanke, D. H.; Carpenter, K. (eds.). Mesozoic Vertebrate Life. Life of the Past. Indiana University Press. pp. 58–63.
- Steven E. Jasinski (2015). "A new dromaeosaurid (Theropoda: Dromaeosauridae) from the Late Cretaceous of New Mexico". New Mexico Museum of Natural History and Science Bulletin. 67: 79–88.
- Longrich, N.R.; Currie, P.J. (2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". Proceedings of the National Academy of Sciences. 106 (13): 5002–5007. Bibcode:2009PNAS..106.5002L. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- Junchang Lü & Stephen L. Brusatte (2015). "A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution". Scientific Reports. 5: Article number 11775. Bibcode:2015NatSR...511775L. doi:10.1038/srep11775. PMC 4504142. PMID 26181289.
- Lü, J.-C.; Xu, L.; Zhang, X.-L.; Ji, Q.; Jia, S.-H.; Hu, W.-Y.; Zhang, J.-M.; Wu, Y.-H. (2007). "New dromaeosaurid dinosaur from the Late Cretaceous Qiupa Formation of Luanchuan area, western Henan, China". Geological Bulletin of China. 26 (7): 777–786.
- Makovicky, Peter J.; Norell, Mark A. (2004). "Troodontidae". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 184–195. ISBN 0-520-24209-2.
- Makovicky, Peter J.; Apesteguía, Sebastián; Agnolín, Federico L. (2005). "The earliest dromaeosaurid theropod from South America". Nature. 437 (7061): 1007–1011. Bibcode:2005Natur.437.1007M. doi:10.1038/nature03996. PMID 16222297. S2CID 27078534.
- Mayor, Adrienne (2005). Fossil Legends of the First Americans. Princeton University Press. ISBN 0-691-11345-9.
- Norell, M.A. & Makovicky, P.J. (2004). "Dromaeosauridae". In Weishampel, D.B.; Dodson, P. & Osmólska, H. (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 196–210. ISBN 0-520-24209-2.
- Norell, M.A.; Clark, J.M.; Turner, A.H.; Makovicky, P.J.; Barsbold, R.; Rowe, T. (2006). "A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)". American Museum Novitates. 3545: 1–51. doi:10.1206/0003-0082(2006)3545[1:andtfu]2.0.co;2. hdl:2246/5823.
- Novas, Fernando E.; Pol, Diego; Canale, Juan I.; Porfiri, Juan D.; Calvo, Jorge O. (2008). "A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids". Proceedings of the Royal Society B. 276 (1659): 1101–1107. doi:10.1098/rspb.2008.1554. PMC 2679073. PMID 19129109.
- Novas, Fernando E.; Pol, Diego (2005). "New Evidence on Deinonychosaurian Dinosaurs from the Late Cretaceous of Patagonia". Nature. 433 (7028): 858–861. Bibcode:2005Natur.433..858N. doi:10.1038/nature03285. PMID 15729340. S2CID 4413316.
- Ostrom, John H. (1969). "Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana". Peabody Museum of Natural History Bulletin. 30: 1–165.
- Padian, K.; Ji, Qiang; Ji, Shu-An (2001). "Feathered dinosaurs and origin of flight". In Tanke, D. H.; Carpenter, K. (eds.). Mesozoic Vertebrate Life. Life of the Past. Indiana University Press. pp. 117–135.
- Porfiri, Juan D.; Jorge O. Calvo; Domenica dos Santos (2011). "A new small deinonychosaur (Dinosauria: Theropoda) from the Late Cretaceous of Patagonia, Argentina" (PDF). Anais da Academia Brasileira de Ciências. 83 (1): 109–116. doi:10.1590/S0001-37652011000100007. ISSN 0001-3765. PMID 21437378.
- Senter, P.; Kirkland, J. I.; Deblieux, D. D.; Madsen, S.; Toth, N. (2012). Dodson, Peter (ed.). "New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail". PLOS ONE. 7 (5): e36790. Bibcode:2012PLoSO...736790S. doi:10.1371/journal.pone.0036790. PMC 3352940. PMID 22615813.
- Turner, A.S.; Hwang, S.H.; Norell, M.A. (2007). "A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia" (PDF). American Museum Novitates. 3557 (1): 1–27. doi:10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2. hdl:2246/5845. Retrieved 2007-03-29.
- Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, Mark (2007). "A basal dromaeosaurid and size evolution preceding avian flight" (PDF). Science. 317 (5843): 1378–1381. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350. S2CID 2519726.
- Xu Xing, Choinere, J., Pittman, M., Tan, Q., Xiao, D., Li, Z., Tan, L., Clark, J., Norell, M., Hone, D.W.E. and Sullivan, C. (19 March 2010). "A new dromaeosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Wulansuhai Formation of Inner Mongolia, China" (PDF). Zootaxa (2403): 1–9. Retrieved 2010-03-19.CS1 maint: uses authors parameter (link)
- Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F.; Du, X. (January 23, 2003). "Four-winged dinosaurs from China". Nature. 421 (6921): 335–340. Bibcode:2003Natur.421..335X. doi:10.1038/nature01342. PMID 12540892. S2CID 1160118.
- Xu, X.; Wang, X.-L. (2004). "A New Dromaeosaur (Dinosauria: Theropoda) from the Early Cretaceous Yixian Formation of Western Liaoning". Vertebrata PalAsiatica. 42 (2): 11–119.
- Zheng, Xiaoting; Xu, Xing; You, Hailu; Zhao, Qi; Dong, Zhiming (2010). "A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution". Proceedings of the Royal Society B. 277 (1679): 211–217. doi:10.1098/rspb.2009.1178. PMC 2842677. PMID 19692406.
External links
 Media related to Dromaeosauridae at Wikimedia Commons
Media related to Dromaeosauridae at Wikimedia Commons