Mothers against decapentaplegic homolog 3
Mothers against decapentaplegic homolog 3 also known as SMAD family member 3 or SMAD3 is a protein that in humans is encoded by the SMAD3 gene.[5][6]
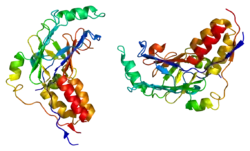




SMAD3 is a member of the SMAD family of proteins. It acts as a mediator of the signals initiated by the transforming growth factor beta (TGF-β) superfamily of cytokines, which regulate cell proliferation, differentiation and death.[7] [8] Based on its essential role in TGF beta signaling pathway, SMAD3 has been related with tumor growth in cancer development.
Gene
The human SMAD3 gene is located on chromosome 15 on the cytogenic band at 15q22.33. The gene is composed of 9 exons over 129,339 base pairs.[9] It is one of several human homologues of a gene that was originally discovered in the fruit fly Drosophila melanogaster.
The expression of SMAD3 has been related to the mitogen-activated protein kinase (MAPK/ERK pathway), particularly to the activity of mitogen-activated protein kinase kinase-1 (MEK1).[10] Studies have demonstrated that inhibition of MEK1 activity also inhibits SMAD3 expression in epithelial cells and smooth muscle cells, two cell types highly responsive to TGF-β1.[10]
Protein
SMAD3 is a polypeptide with a molecular weight of 48,080 Da. It belongs to the SMAD family of proteins. SMAD3 is recruited by SARA (SMAD Anchor for Receptor Activation) to the membrane, where the TGF-β receptor is located. The receptors for TGF-β, (including nodal, activin, myostatin and other family members) are membrane serine/threonine kinases that preferentially phosphorylate and activate SMAD2 and SMAD3.
Once SMAD3 is phosphorylated at the C-terminus, it dissociates from SARA and forms a heterodimeric complex with SMAD4, which is required for the transcriptional regulation of many target genes.[11] The complex of two SMAD3 (or of two SMAD2) and one SMAD4 binds directly to DNA though interactions of the MH1 domain. These complexes are recruited to sites throughout the genome by cell lineage-defining transcription factors (LDTFs) that determine the context-dependent nature of TGF-β action. The DNA binding sites in promoters and enhancers are known as the SMAD-binding elements (SBEs). These sites contain the CAG(AC)|(CC) and GGC(GC)|(CG) consensus sequences, the latter also known as 5GC sites.[12] The 5GC-motifs are highly represented as clusters of sites, in SMAD-bound regions genome-wide. These clusters can also contain CAG(AC)|(CC) sites. SMAD3/SMAD4 complex also binds to the TPA-responsive gene promoter elements, which have the sequence motif TGAGTCAG.[13]
Structure
MH1 domain
The X-ray structures of the SMAD3 MH1 domain bound to the GTCT DNA reveal characteristic features of the fold. The MH1 structure consists of four-helices and three sets of antiparallel β-hairpins, one of which is used to interact with DNA. It also revealed the presence of a bound Zn2+, coordinated by His126, Cys64, Cys109 and Cys121 residues.[11][12] The main DNA binding region of the MH1 domain comprises the loop following the β1 strand, and the β2-β3 hairpin. In the complex with a member of the 5GC DNAs, the GGCGC motif, the convex face of the DNA-binding hairpin dives into the concave major groove of the duplex DNA containing five base pairs (GGCGC /'GCGCC'). In addition, the three residues strictly conserved in all R-SMADS and in SMAD4 (Arg74 and Gln76 located in β2 and Lys81 in β3 in SMAD3) participate in a network of specific hydrogen bonds with the dsDNA. Several tightly bound water molecules at the protein-DNA interface that contribute to the stabilization of the interactions have also been detected. The SMAD3 complex with the GGCGC site reveals that the protein-DNA interface is highly complementary and that one MH1 protein covers a DNA binding site of six base pairs.
MH2 domain
The MH2 domain mediates the interaction of R-SMADS with activated TGF-β receptors, and with SMAD4 after receptor-mediated phosphorylation of the Ser-X-Ser motif present in R-SMADS. The MH2 domain is also a binding platform for cytoplasmic anchors, DNA-binding cofactors, histone modifiers, chromatin readers, and nucleosome- positioning factors. The structure of the complex of SMAD3 and SMAD4 MH2 domains has been determined.[14] The MH2 fold is defined by two sets of antiparallel β-strands (six and five strands respectively) arranged as a β-sandwich flanked by a triple-helical bundle on one side and by a set of large loops and a helix on the other.
Functions and interactions
TGF-β/SMAD signaling pathway
SMAD3 functions as a transcriptional modulator, binding the TRE (TPA responsive element) in the promoter region of many genes that are regulated by TGF-β. SMAD3 and SMAD4 can also form a complex with c-Fos and c-jun at the AP-1/SMAD site to regulate TGF-β-inducible transcription.[13] The genes regulated by SMAD3-mediated TGFβ signaling affect differentiation, growth and death. TGF-β/SMAD signaling pathway has been shown to have a critical role in the expression of genes controlling differentiation of embryonic stem cells.[15] Some of the developmental genes regulated by this pathway include FGF1, NGF, and WNT11 as well as stem/progenitor cell associated genes CD34 and CXCR4.[16] The activity of this pathway as a regulator of pluripotent cell states requires the TRIM33-SMAD2/3 chromatin reading complex.[15]
TGF-β/SMAD3-induced repression
Besides the activity of TGF-β in the up-regulation of genes, this signaling molecule also induces the repression of target genes containing the TGF-β inhibitory element (TIE).[17][18] SMAD3 plays also a critical role in TGF-β-induced repression of target genes, specifically it is required for the repression of c-myc.The transcriptional repression of c-myc is dependent on direct SMAD3 binding to a repressive SMAD binding element (RSBE), within TIE of the c-myc promoter. The c-myc TIE is a composite element, composed of an overlapping RSBE and a consensus E2F site, which is capable of binding at least SMAD3, SMAD4, E2F4, and p107.[18]
Clinical significance
Diseases
Increased SMAD3 activity has, however, been implicated in the pathogenesis of scleroderma.
SMAD3 is also a multifaceted regulator in adipose physiology and the pathogenesis of obesity and type 2 diabetes. SMAD3-knockout mice have diminished adiposity,[19] with improved glucose tolerance and insulin sensitivity. Despite their reduced physical activity arising from muscle atrophy,[20] these SMAD3-knockout mice are resistant to high-fat-diet induced obesity. SMAD3-knockout mouse is a legitimate animal model of human aneurysms‐osteoarthritis syndrome(AOS), also named Loeys-Dietz Syndrome (type 3). SMAD3 deficiency promotes inflammatory aortic aneurysms in angiotensin II-infused mice via the activation of iNOS. Macrophage depletion and inhibition of iNOS activity prevent aortic aneurysms related to SMAD3 gene mutation[21]
Role in cancer
The role of SMAD3 in the regulation of genes important for cell fate, such as differentiation, growth and death, implies that an alteration in its activity or repressing of its activity can lead to the formation or development of cancer. Also several studies have proven the bifunctional tumor suppressor/oncogene role of TGF beta signaling pathway in carcinogenesis.[22]
One way in which SMAD3 transcriptional activator function is repressed, is by the activity of EVI-1.[23] EVI-1 encodes a zinc-finger protein that may be involved in leukaemic transformation of haematopoietic cells. The zinc-finger domain of EVI-1 interacts with SMAD3, thereby suppressing the transcriptional activity of SMAD3. EVI-1 is thought to be able to promote growth and to block differentiation in some cell types by repressing TGF-β signalling and antagonizing the growth-inhibitory effects of TGF-β.[23]
Prostate
The activity of SMAD3 in prostate cancer is related with the regulation of angiogenic molecules expression in tumor vascularization and cell-cycle inhibitor in tumor growth.[24][25] The progressive growth of primary tumors and metastases in prostate cancer depends on an adequate blood supply provided by tumor angiogenesis. Studies analyzing SMAD3 levels of expression in prostate cancer cell lines found that the two androgen-independent and androgen receptor-negative cell lines (PC-3MM2 and DU145) have high expression levels of SMAD3. Analysis of the relation between SMAD3 and the regulation of angiogenic molecules suggest that SMAD3 may be one of key components as a repressor of the critical angiogenesis switch in prostate cancer.[25] The pituitary tumor-transforming gene 1 (PTTG1) has also an impact in SMAD3-mediated TGFβ signaling. PTTG1 has been associated with various cancer cells including prostate cancer cells. Studies showed that the overexpression of PTTG1 induces a decrease in SMAD3 expression, promoting the proliferation of prostate cancer cells via the inhibition of SMAD3.[24]
Colorectal
In mice, mutation of SMAD3 has been linked to colorectal adenocarcinoma,[3] increased systemic inflammation, and accelerated wound healing.[4] Studies have shown that mutations in SMAD3 gene promote colorectal cancer in mice.[26][27][28] The altered activity of SMAD3 was linked to chronic inflammation and somatic mutations that contribute to chronic colitis and the development of colorectal cancer.[28] The results generated on mice helped identify SMAD3 like a possible player in human colorectal cancer. The impact of SMAD3 has also been analyzed in colorectal cancer human cell lines, using single-nucleotide polymorphism (SNP) microarray analysis. The results showed reductions in SMAD3 transcriptional activity and SMAD2-SMAD4 complex formation, underlining the critical roles of these three proteins within the TGF-β signaling pathway and the impact of this pathway in colorectal cancer development.[29]
Breast
TGF-β-induced SMAD3 transcriptional regulation response has been associated with breast cancer bone metastasis by its effects on tumor angiogenesis, and epithelial-mesenchymal transition (EMT). There have been identified diverse molecules that act over the TGF-β/SMAD signaling pathway, affecting primarily the SMAD2/3 complex, which have been associated with the development of breast cancer.[30]
FOXM1 (forkhead box M1) is a molecule that binds with SMAD3 to sustain activation of the SMAD3/SMAD4 complex in the nucleus. The research over FOXM1 suggested that it prevents the E3 ubiquitin-protein ligase transcriptional intermediary factor 1 γ (TIF1γ) from binding SMAD3 and monoubiquitinating SMAD4, which stabilized the SMAD3/SMAD4 complex. FOXM1 is a key player in the activity of the SMAD3/SMAD4 complex, promoting SMAD3 modulator transcriptional activity, and also plays an important role in the turnover of the activity of SMAD3/SMAD4 complex. Based on the importance of this molecule, studies have found that FOXM1 is overexpressed in highly aggressive human breast cancer tissues. The results from these studies also found that the FOXM1/SMAD3 interaction was required for TGF-β-induced breast cancer invasion, which was the result of SMAD3/SMAD4-dependent upregulation of the transcription factor SLUG.[31]
MED15 is a mediator molecule that promotes the activity of the TGF-β/SMAD signaling. The deficiency of this molecule attenuates the activity of the TGF-β/SMAD signaling pathway over the genes required for induction of epithelial-mesenchymal transition. The action of MED15 is related with the phosphorylation of SMAD2/3 complex. The knockdown of MED15 reduces the amount of SMAD3 phosphorylated, therefore reducing its activity as transcription modulator. However, in cancer, MED15 is also highly expressed in clinical breast cancer tissues correlated with hyperactive TGF-β signaling, as indicated by SMAD3 phosphorylation. The studies suggest that MED15 increases the metastatic potential of a breast cancer cell line by increasing TGF-β-induced epithelial–mesenchymal transition.[32]
Kidney
Smad3 activation plays a role in the pathogenesis of renal fibrosis,[33] probably by inducing activation of bone marrow-derived fibroblasts [34]
Nomenclature
The SMAD proteins are homologs of both the Drosophila protein "mothers against decapentaplegic" (MAD) and the C. elegans protein SMA. The name is a combination of the two. During Drosophila research, it was found that a mutation in the gene MAD in the mother repressed the gene decapentaplegic in the embryo. The phrase "Mothers against" was inspired by organizations formed by mothers to oppose social problems, such as Mothers Against Drunk Driving (MADD); and based on a tradition of such unusual naming within the gene research community.[35]
A reference assembly of SMAD3 is available.
References
- GRCh38: Ensembl release 89: ENSG00000166949 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000032402 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: SMAD3 SMAD family member 3".
- Zhang Y, Feng X, We R, Derynck R (September 1996). "Receptor-associated Mad homologues synergize as effectors of the TGF-beta response". Nature. 383 (6596): 168–72. Bibcode:1996Natur.383..168Z. doi:10.1038/383168a0. PMID 8774881. S2CID 4306019.
- Massagué J (1998). "TGF-beta signal transduction". Annual Review of Biochemistry. 67 (1): 753–91. doi:10.1146/annurev.biochem.67.1.753. PMID 9759503.
- Moustakas A, Souchelnytskyi S, Heldin CH (December 2001). "Smad regulation in TGF-beta signal transduction". Journal of Cell Science. 114 (Pt 24): 4359–69. PMID 11792802.
- GeneCards. "SMAD3 Gene".
- Ross KR, Corey DA, Dunn JM, Kelley TJ (May 2007). "SMAD3 expression is regulated by mitogen-activated protein kinase kinase-1 in epithelial and smooth muscle cells". Cellular Signalling. 19 (5): 923–31. doi:10.1016/j.cellsig.2006.11.008. PMID 17197157.
- Shi, Yigong; Massagué, Joan (2003). "Mechanisms of TGF-β Signaling from Cell Membrane to the Nucleus". Cell. 113 (6): 685–700. doi:10.1016/S0092-8674(03)00432-X. ISSN 0092-8674. PMID 12809600. S2CID 16860578.
- Martin-Malpartida P, Batet M, Kaczmarska Z, Freier R, Gomes T, Aragón E, Zou Y, Wang Q, Xi Q, Ruiz L, Vea A, Márquez JA, Massagué J, Macias MJ (December 2017). "Structural basis for genome wide recognition of 5-bp GC motifs by SMAD transcription factors". Nature Communications. 8 (1): 2070. Bibcode:2017NatCo...8.2070M. doi:10.1038/s41467-017-02054-6. PMC 5727232. PMID 29234012.
- Zhang Y, Feng XH, Derynck R (August 1998). "Smad3 and Smad4 cooperate with c-Jun/c-Fos to mediate TGF-beta-induced transcription". Nature. 394 (6696): 909–13. Bibcode:1998Natur.394..909Z. doi:10.1038/29814. PMID 9732876. S2CID 4393852.
- Chacko BM, Qin BY, Tiwari A, Shi G, Lam S, Hayward LJ, De Caestecker M, Lin K (September 2004). "Structural basis of heteromeric smad protein assembly in TGF-beta signaling". Molecular Cell. 15 (5): 813–23. doi:10.1016/j.molcel.2004.07.016. PMID 15350224.
- Massagué J, Xi Q (July 2012). "TGF-β control of stem cell differentiation genes". FEBS Letters. 586 (14): 1953–8. doi:10.1016/j.febslet.2012.03.023. PMC 3466472. PMID 22710171.
- Shi X, DiRenzo D, Guo LW, Franco SR, Wang B, Seedial S, Kent KC (2014). "TGF-β/Smad3 stimulates stem cell/developmental gene expression and vascular smooth muscle cell de-differentiation". PLOS ONE. 9 (4): e93995. Bibcode:2014PLoSO...993995S. doi:10.1371/journal.pone.0093995. PMC 3981734. PMID 24718260.
- Matrisian LM, Ganser GL, Kerr LD, Pelton RW, Wood LD (June 1992). "Negative regulation of gene expression by TGF-beta". Molecular Reproduction and Development. 32 (2): 111–20. doi:10.1002/mrd.1080320206. PMID 1637549.
- Frederick JP, Liberati NT, Waddell DS, Shi Y, Wang XF (March 2004). "Transforming growth factor beta-mediated transcriptional repression of c-myc is dependent on direct binding of Smad3 to a novel repressive Smad binding element". Molecular and Cellular Biology. 24 (6): 2546–59. doi:10.1128/mcb.24.6.2546-2559.2004. PMC 355825. PMID 14993291.
- Tan CK, Leuenberger N, Tan MJ, Yan YW, Chen Y, Kambadur R, Wahli W, Tan NS (February 2011). "Smad3 deficiency in mice protects against insulin resistance and obesity induced by a high-fat diet". Diabetes. 60 (2): 464–76. doi:10.2337/db10-0801. PMC 3028346. PMID 21270259.
- Ge X, McFarlane C, Vajjala A, Lokireddy S, Ng ZH, Tan CK, Tan NS, Wahli W, Sharma M, Kambadur R (November 2011). "Smad3 signaling is required for satellite cell function and myogenic differentiation of myoblasts". Cell Research. 21 (11): 1591–604. doi:10.1038/cr.2011.72. PMC 3364732. PMID 21502976.
- Tan CK, Tan EH, Luo B, Huang CL, Loo JS, Choong C, Tan NS (June 2013). "SMAD3 deficiency promotes inflammatory aortic aneurysms in angiotensin II-infused mice via activation of iNOS". Journal of the American Heart Association. 2 (3): e000269. doi:10.1161/JAHA.113.000269. PMC 3698794. PMID 23782924.
- de Caestecker MP, Piek E, Roberts AB (September 2000). "Role of transforming growth factor-beta signaling in cancer". Journal of the National Cancer Institute. 92 (17): 1388–402. doi:10.1093/jnci/92.17.1388. PMID 10974075.
- Kurokawa M, Mitani K, Irie K, Matsuyama T, Takahashi T, Chiba S, Yazaki Y, Matsumoto K, Hirai H (July 1998). "The oncoprotein Evi-1 represses TGF-beta signalling by inhibiting Smad3". Nature. 394 (6688): 92–6. Bibcode:1998Natur.394...92K. doi:10.1038/27945. PMID 9665135. S2CID 4404132.
- Huang S, Liao Q, Li L, Xin D (July 2014). "PTTG1 inhibits SMAD3 in prostate cancer cells to promote their proliferation". Tumour Biology. 35 (7): 6265–70. doi:10.1007/s13277-014-1818-z. PMID 24627133. S2CID 17153729.
- Lu S, Lee J, Revelo M, Wang X, Lu S, Dong Z (October 2007). "Smad3 is overexpressed in advanced human prostate cancer and necessary for progressive growth of prostate cancer cells in nude mice". Clinical Cancer Research. 13 (19): 5692–702. doi:10.1158/1078-0432.CCR-07-1078. PMID 17908958. S2CID 14496617.
- Hachimine D, Uchida K, Asada M, Nishio A, Kawamata S, Sekimoto G, Murata M, Yamagata H, Yoshida K, Mori S, Tahashi Y, Matsuzaki K, Okazaki K (June 2008). "Involvement of Smad3 phosphoisoform-mediated signaling in the development of colonic cancer in IL-10-deficient mice". International Journal of Oncology. 32 (6): 1221–6. doi:10.3892/ijo.32.6.1221. PMID 18497983.
- Seamons A, Treuting PM, Brabb T, Maggio-Price L (2013). "Characterization of dextran sodium sulfate-induced inflammation and colonic tumorigenesis in Smad3(-/-) mice with dysregulated TGFβ". PLOS ONE. 8 (11): e79182. Bibcode:2013PLoSO...879182S. doi:10.1371/journal.pone.0079182. PMC 3823566. PMID 24244446.
- Kawamata S, Matsuzaki K, Murata M, Seki T, Matsuoka K, Iwao Y, Hibi T, Okazaki K (March 2011). "Oncogenic Smad3 signaling induced by chronic inflammation is an early event in ulcerative colitis-associated carcinogenesis". Inflammatory Bowel Diseases. 17 (3): 683–95. doi:10.1002/ibd.21395. PMID 20602465. S2CID 5136295.
- Fleming NI, Jorissen RN, Mouradov D, Christie M, Sakthianandeswaren A, Palmieri M, Day F, Li S, Tsui C, Lipton L, Desai J, Jones IT, McLaughlin S, Ward RL, Hawkins NJ, Ruszkiewicz AR, Moore J, Zhu HJ, Mariadason JM, Burgess AW, Busam D, Zhao Q, Strausberg RL, Gibbs P, Sieber OM (January 2013). "SMAD2, SMAD3 and SMAD4 mutations in colorectal cancer". Cancer Research. 73 (2): 725–35. doi:10.1158/0008-5472.CAN-12-2706. PMID 23139211.
- Petersen M, Pardali E, van der Horst G, Cheung H, van den Hoogen C, van der Pluijm G, Ten Dijke P (March 2010). "Smad2 and Smad3 have opposing roles in breast cancer bone metastasis by differentially affecting tumor angiogenesis". Oncogene. 29 (9): 1351–61. doi:10.1038/onc.2009.426. PMID 20010874. S2CID 11592749.
- Xue J, Lin X, Chiu WT, Chen YH, Yu G, Liu M, Feng XH, Sawaya R, Medema RH, Hung MC, Huang S (February 2014). "Sustained activation of SMAD3/SMAD4 by FOXM1 promotes TGF-β-dependent cancer metastasis". The Journal of Clinical Investigation. 124 (2): 564–79. doi:10.1172/JCI71104. PMC 3904622. PMID 24382352.
- Zhao M, Yang X, Fu Y, Wang H, Ning Y, Yan J, Chen YG, Wang G (February 2013). "Mediator MED15 modulates transforming growth factor beta (TGFβ)/Smad signaling and breast cancer cell metastasis". Journal of Molecular Cell Biology. 5 (1): 57–60. doi:10.1093/jmcb/mjs054. PMID 23014762.
- Meng XM, Chung AC, Lan HY (February 2013). "Role of the TGF-β/BMP-7/Smad pathways in renal diseases". Clinical Science. 124 (4): 243–54. doi:10.1042/CS20120252. PMID 23126427.
- Chen J, Xia Y, Lin X, Feng XH, Wang Y (May 2014). "Smad3 signaling activates bone marrow-derived fibroblasts in renal fibrosis". Laboratory Investigation; A Journal of Technical Methods and Pathology. 94 (5): 545–56. doi:10.1038/labinvest.2014.43. PMC 4006302. PMID 24614197.
- "Sonic Hedgehog, DICER, and the Problem With Naming Genes", Sep 26, 2014, Michael White. psmag.com
Further reading
- Tan CK, Chong HC, Tan EH, Tan NS (March 2012). "Getting 'Smad' about obesity and diabetes". Nutrition & Diabetes. 2 (3): e29. doi:10.1038/nutd.2012.1. PMC 3341711. PMID 23449528.
- Li H, Liu JP (October 2007). "Mechanisms of action of TGF-beta in cancer: evidence for Smad3 as a repressor of the hTERT gene". Annals of the New York Academy of Sciences. 1114 (1): 56–68. Bibcode:2007NYASA1114...56L. doi:10.1196/annals.1396.016. PMID 17934056.
- Matsuzaki K (June 2006). "Smad3 phosphoisoform-mediated signaling during sporadic human colorectal carcinogenesis". Histology and Histopathology. 21 (6): 645–62. doi:10.14670/HH-21.645. PMID 16528675.
- Miyazono K (2000). "TGF-beta signaling by Smad proteins". Cytokine & Growth Factor Reviews. 11 (1–2): 15–22. doi:10.1016/S1359-6101(99)00025-8. PMID 10708949.
- Wrana JL, Attisano L (2000). "The Smad pathway". Cytokine & Growth Factor Reviews. 11 (1–2): 5–13. doi:10.1016/S1359-6101(99)00024-6. PMID 10708948.
- Verschueren K, Huylebroeck D (2000). "Remarkable versatility of Smad proteins in the nucleus of transforming growth factor-beta activated cells". Cytokine & Growth Factor Reviews. 10 (3–4): 187–99. doi:10.1016/S1359-6101(99)00012-X. PMID 10647776.
- Massagué J (1998). "TGF-beta signal transduction". Annual Review of Biochemistry. 67: 753–91. doi:10.1146/annurev.biochem.67.1.753. PMID 9759503.
- Walker LC, Waddell N, Ten Haaf A, Grimmond S, Spurdle AB (November 2008). "Use of expression data and the CGEMS genome-wide breast cancer association study to identify genes that may modify risk in BRCA1/2 mutation carriers". Breast Cancer Research and Treatment. 112 (2): 229–36. doi:10.1007/s10549-007-9848-5. PMID 18095154. S2CID 795870.
- Lee KB, Jeon JH, Choi I, Kwon OY, Yu K, You KH (February 2008). "Clusterin, a novel modulator of TGF-beta signaling, is involved in Smad2/3 stability". Biochemical and Biophysical Research Communications. 366 (4): 905–9. doi:10.1016/j.bbrc.2007.12.033. PMID 18082619.
- Kim TD, Shin S, Janknecht R (February 2008). "Repression of Smad3 activity by histone demethylase SMCX/JARID1C". Biochemical and Biophysical Research Communications. 366 (2): 563–7. doi:10.1016/j.bbrc.2007.12.013. PMID 18078810.
- Zhao X, Nicholls JM, Chen YG (February 2008). "Severe acute respiratory syndrome-associated coronavirus nucleocapsid protein interacts with Smad3 and modulates transforming growth factor-beta signaling". The Journal of Biological Chemistry. 283 (6): 3272–80. doi:10.1074/jbc.M708033200. PMID 18055455. S2CID 84455415.
- Li T, Chiang JY (November 2007). "A novel role of transforming growth factor beta1 in transcriptional repression of human cholesterol 7alpha-hydroxylase gene". Gastroenterology. 133 (5): 1660–9. doi:10.1053/j.gastro.2007.08.042. PMID 17920062.
- Lu S, Lee J, Revelo M, Wang X, Lu S, Dong Z (October 2007). "Smad3 is overexpressed in advanced human prostate cancer and necessary for progressive growth of prostate cancer cells in nude mice". Clinical Cancer Research. 13 (19): 5692–702. doi:10.1158/1078-0432.CCR-07-1078. PMID 17908958. S2CID 14496617.
- Kalo E, Buganim Y, Shapira KE, Besserglick H, Goldfinger N, Weisz L, Stambolsky P, Henis YI, Rotter V (December 2007). "Mutant p53 attenuates the SMAD-dependent transforming growth factor beta1 (TGF-beta1) signaling pathway by repressing the expression of TGF-beta receptor type II". Molecular and Cellular Biology. 27 (23): 8228–42. doi:10.1128/MCB.00374-07. PMC 2169171. PMID 17875924.
- Weng HL, Ciuclan L, Liu Y, Hamzavi J, Godoy P, Gaitantzi H, Kanzler S, Heuchel R, Ueberham U, Gebhardt R, Breitkopf K, Dooley S (October 2007). "Profibrogenic transforming growth factor-beta/activin receptor-like kinase 5 signaling via connective tissue growth factor expression in hepatocytes". Hepatology. 46 (4): 1257–70. doi:10.1002/hep.21806. PMID 17657819.
- Dennler S, André J, Alexaki I, Li A, Magnaldo T, ten Dijke P, Wang XJ, Verrecchia F, Mauviel A (July 2007). "Induction of sonic hedgehog mediators by transforming growth factor-beta: Smad3-dependent activation of Gli2 and Gli1 expression in vitro and in vivo". Cancer Research. 67 (14): 6981–6. doi:10.1158/0008-5472.CAN-07-0491. PMID 17638910.
- Zhang M, Lee CH, Luo DD, Krupa A, Fraser D, Phillips A (September 2007). "Polarity of response to transforming growth factor-beta1 in proximal tubular epithelial cells is regulated by beta-catenin". The Journal of Biological Chemistry. 282 (39): 28639–47. doi:10.1074/jbc.M700594200. PMID 17623674. S2CID 41791801.
- Martin MM, Buckenberger JA, Jiang J, Malana GE, Knoell DL, Feldman DS, Elton TS (September 2007). "TGF-beta1 stimulates human AT1 receptor expression in lung fibroblasts by cross talk between the Smad, p38 MAPK, JNK, and PI3K signaling pathways". American Journal of Physiology. Lung Cellular and Molecular Physiology. 293 (3): L790-9. doi:10.1152/ajplung.00099.2007. PMC 2413071. PMID 17601799.
- Dai F, Chang C, Lin X, Dai P, Mei L, Feng XH (September 2007). "Erbin inhibits transforming growth factor beta signaling through a novel Smad-interacting domain". Molecular and Cellular Biology. 27 (17): 6183–94. doi:10.1128/MCB.00132-07. PMC 1952163. PMID 17591701.
- Levy L, Howell M, Das D, Harkin S, Episkopou V, Hill CS (September 2007). "Arkadia activates Smad3/Smad4-dependent transcription by triggering signal-induced SnoN degradation". Molecular and Cellular Biology. 27 (17): 6068–83. doi:10.1128/MCB.00664-07. PMC 1952153. PMID 17591695.